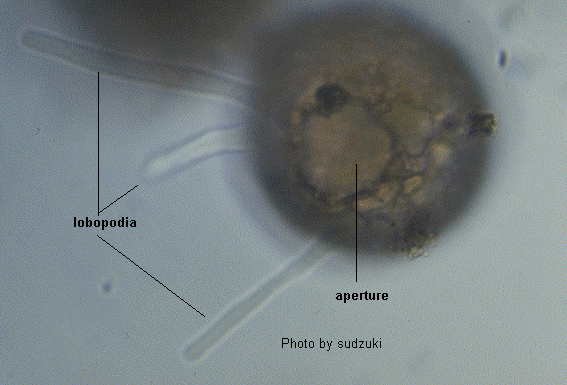
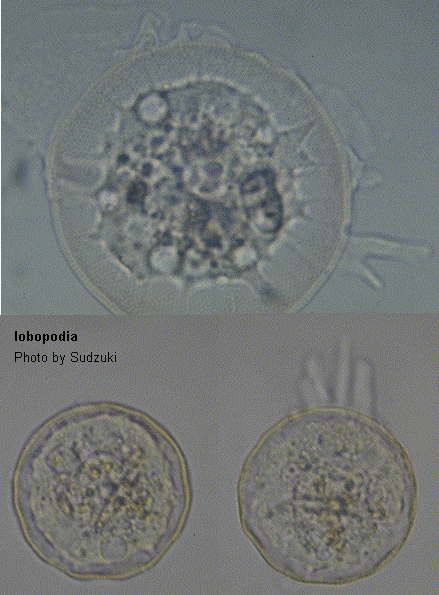
[Notice]
We reproduce the descriptions of morphology from following books;
ref. ID; 1618
Kudo, "Protozoology 5th Ed."
Copyright: CHARLES C THOMAS PUBLISHER, LTD., 1966
ref. ID; 3686
C.G. Ogden and R.H. Hedley, "An Atlas of Freshwater Testate Amoebae"
ISBN 0 19 858502 0
Copyright: The Natural History Museum, London
On-line catalogue available at: http://www.nhm.ac.uk/services/publishing/
On-line bookshop at: http://www.nhm.ac.uk/shop/index.html
and we thank following scientists and The Society of Protozoologists (The Journal of Protozoology) for quoting text descriptions from their papers.
Review on the Variability of Testate Amoebae [ref. ID; 2852 (Wanner 1999)]
Testate amoebae are particularly suitable for the fundamental question, whether and how an organism responds to changing environments. They respond to modified conditions by altering abundance or dominance structure (Lousier 1974; Foissner 1987, 1997) and by changing their shell morphometry (e.g. Wanner and Meisterfeld 1994; Wanner 1994, 1995). The shell architecture of testate amoebae (e.g. shell type and shape, spikes, spines, diaphragms, the aperture) has been commonly used of differentiate between genera or species. Physical limitations, like diffusion-dependent cells size range, surface tension, or biomechanical preconditions of protists skeletons (see Rhumbler 1898; Vogel and Gutmann 1988; Fenchel 1990) and ecological constraints, like habitat adaptation, will also influence the shape of the amoebae shell, resulting in a great variety of different morphs. For taxonomical and ecological reasons it is important to estimate the range and form ("genetic", "nongenetic", as defined in Mayr 1969) of variability within a given taxon. As stressed in Mayr (1969), "the underestimation of individual variation may have caused more than 50% of all synonyms". Furthermore, he stated that "differences between groups of similar specimens ("phena") may reflect either a speices difference or intraspecific variation. Therefore a complete understanding of intraspecific variation is necessary before we are able to separate between possible species." Is for instance the phenomenon of ecomorphosis, the variation caused by the environment (expressed by changes in shell morphology) not only measurable, but also reproducible or even reversible within a few generations? If so, this would give new and fascinating tools for bioindication with testate amoebae, in the laboratory as well as in the field. On the other hand, a strong influence of environmental factors on shell morphometry would lead to serious taxonomic problems, because classification of closely related testate amoebae is primarily based on these characteristics.
General features of testate amoebae
The shell encloses the cell plasma and has usually a single aperture for the pseudopodia. A poteinaceous organic matrix is the basic shell component, either solely or functioning as cement, fixing particles in position (Moraczewski 1969; Saucin-Meulenberg et al. 1973). There are four main shell types: proteinaceous (species with a flexible or rigit shell), calcareous (only two genera), siliceous (species which secrete their own regular siliceous shell platelets, so-called "idiosomes"), and agglutinate (species which include extraneous mineral particles in their shell structure, so-called "xenosomes"). Details are discussed by Grospietsch (1958), Schonborn (1966), Netzel (1983), Ogden and Hedley (1980), Ogden (1984, 1990, 1991), Anderson (1987), and Foissner (1987). Testate amoebae are a polyphyletic, or at least biphyletic, assemblage. The major characteristics, the shell and the pseudopodia, evolved independently representing only convergent features (Hausmann and Hulsmann 1996). This view is strongly supported by molecular data (Bhattacharya et al. 1995; Cavalier-Smith 1997). At present, naked and testate filose amoebae are grouped within the (revised) phylum Rhizopoda, while naked and testate lobose amoebae are grouped into a separate (revised) sarcodine phylum, the Amoebozoa (Cavalier-Smith 1997). This is supported by community structure and biometric data. Foissner (1987) and Wodarz et al. (1992) pointed out that taxa of different origin (e.g. lobose and filose testates) may have a different autecology with individual importance for bioindication. Based on a biometrical analysis of twenty-four soil testate amoebae, Luftenegger et al. (1988) observed that Testaceafilosa have wider ranges in morphometric variation as compared with Testacealobosea.
Methodological Approaches
The basic methodological tools for collecting, preparation, observation and statistics have been discussed in detail by Bonnet (1964), Schonborn (1966), Laminger (1980), Foissner (1987, 1994), Wanner (1991), Aescht and Foissner (1995), and Dunger and Fiedler (1997). Hence in this chapter some specific methodological problems will be highlighted concerning the analysis and interpretation of variabiliy in testate amoebae.
Field investigations. They provide a "natural environment" for experimental and descriptive analyses. An objective, artifact-free assessment of the environmentally depended variability will be possible, including all relevant factors and interactions which are affecting the organism of interest. However, this requires a profound and time-consuming quantitative estimation of the spatial and temporal distribution of the entire important (what is important?) environmental factors using e.g. a completely randomized block design (Hurlbert 1984; Kohler et al. 1984; Petz and Foissner 1989; Wanner et al. 1994). Furthermore, environmental effects acting indirectly are hardly to assess. Thus it is appropriate to combine field data with those from the laboratory.
Laboratory experiments. They allow an unequivocal assessment of single, selected environmental factors - acting directly and indirectly - under standardized are reproducible conditions. However, only if detailed information about the intrinsic intraclonal variability is available (concerning cultures kept under constant conditions), a laboratory estimation of environmental influences and a delimitation of taxa will be successful. For this purpose, numerous taxa of testate amoebae had been isolated, cloned (in this context: cells within a culture descending from one isolated specimen), and successfully cultivated in (batch) cultures for even several years. Quantitative information about the morphological variation is testate amoebae can be attained by measuring different shell parameters (e.g. shell length, - breadth, - depth, apertural size) from about 20-30 specimen (see Hoogenraad and de Groot 1937; Meisterfeld and Schuller 1982; Schonborn et al. 1983). With these biometrical data an "Idealindividuum" was constructed, suitable; to compare species, populations or clones (Schonborn et al. 1983). Significance tests combined with the analysis of size frequency distribution may help to examine geographic races and ecological variations of testate amoebae (Gillies 1918; Luftenegger et al. 1988; Schonborn and Peschke 1988; Schoborn 1990; Luftenegger and Foissner 1991). Precise biometric analysis were also performed using computer image analysis (Wanner 1991), vector-analysis (Meisterfeld and Schuller 1982) and multivariate statistics allowing examinations of numerous clones or populations of the same taxon as well as the quantification of the shell platelets (Wanner 1991, 1995). Cytochemical and X-ray microanalyses of the amoeba shell seem to be further methods for ecological and taxonomical purposes. Stout and Walker (1976) proposed that test of different taxa may be distinguished not only by their size and shape, but also by the chemical character of the mineral particles they are composed of. Although important results concerning the amoeba shell have been revealed by this approach - e.g. it was shown that the ionic compositions of the brown and white clones of Assulina muscorum are nearly identical (Schonborn and Peschke 1990) - definite and consistent conclusions about local differences within populations or between closely related species could not be drawn (e.g. Awerinzew 1907; Couteaux and Jeanson 1977; Rauenbusch 1987). However, further research is necessary to clarify if this tool is suitable for research in amoebal variability.
Molecular methods. Taxon-specific DNA probes may be suitable for the identification and characterization of even closely related species excluding modifying environmental influences. Until recently (Bhattacharya et al. 1995; Wanner et al. 1997), molecular techniques have not been applied on testate amoebae. In most cases neither sufficient amounts of uniform genetical material nor sequence information are available, necessary for RFLP, PCR or sequencing. Additionally, uncommon foreign DNA sources (distinct from the food or the culture medium and located within the amoeba) can inevitably be transferred with the amoebae to each new subculture. This foreign DNA probably belongs to eukaryotic parasites, commensals of symbionts which live inside the amoeba-cell, but are also able to leave the host (Schonborn 1966; Foissner 1987; Anderson and Cowling 1994). As a result the amoeba-free culture medium is contaminated, producing putative "amoebae-clone-specific" DNA patterns (Wanner et al. 1997). This severe problem of coamplifying foreion, but culture-specific DNA, can be prevented by using single nuclei of testate amoebae as a reference. Specific primers derived from a sequenced RAPD-DNA fragment demonstrate its amoeba-specific nuclear origin. These RAPD-derived specific primers allow highly specific amplications of amoebae DNA amplified from a single nucleus, even in the presence of large amounts of contaminating foreign DNA. This approach provides the basic tool for more detailed molecular identification studies in the future (Foissner and Wanner 1995; Wanner et al. 1997). Putative contaminated genetic material of testate amoebae can be checked by a taxon-specific probe. If no eukaryotic contamination is present, an estimation of genetic variation among soil isolated of testate amoebae may be possible, using a combination of the polymerase chain reaction and the restriction fragment length polymorphism analysis (PCR, RFLP), as described of the soil ciliate Colpoda inflata (Bowers and Pratt 1995). But even in the case of eukaryotic contaminations DNA sequence information derived from a few nuclei may be available, which can be clearly assigned to the respective clone of testate amoebae. Fine tuning these methods as well as generating more specific primer combinations should enable reproducible DNA analysis with respect to intraspecific variation, even from small field samples.
Environmental influences
Morphological adaptations. Numerous authors investigated the effects of natural (e.g. moisture, food, temperature, pH, particle size of the habitat) and anthropogenic (e.g. soil compaction, liming, fertilization, pesticides) environmental factors on testate amoebae. Laboratory and field investigations were excellently summarized by Schonborn (1966), and with respect to soil protozoa, by Foissner (1987, 1994, 1997). It was concluded that most morphological adaptations of terrestrial testate amoebae are related to the resticted water resource of the habitat. In general, terrestrial species of the same genus are smaller than the respective freshwater species. Besides moisture, food is also proposed to be an important factor determining the presence or absence of a given species (Martin 1971). Details about the effects of food on intraspecific variability of testate amoebae were presented by Anderson (1989), who worked cultures of Netzelia tuberculata. The gross morphology of the amoebae shell was not significantly changed by the various food items (either yeast or algae). However, the fine structure of the shell platelets was clearly different exhibiting more regular-shaped, smooth platelets in yeast-fed cells compared to those fed on algae. These differences became visible within one generation after transfer to yeast-containing culture medum. Laminger (1978) reported relationships between soil moisture, shell size and kind of food ingested by Trinema enchelys. An increase of shell size was correlated with an increase of soil moisture. Furthermore, the food consumption of Trinema enchelys seemed to vary in relation to soil moisture and shell size. Ogden (1988) firstly reported a significant change in shell architecture (structure and diameter of the aperture) due to agglutinate material in cultures of Cyclopyxis kahli. To conclude, numerous environmental factors do affect shell morphology of testate amoebae in manifold ways, impeding taxonomy which is based on shell size and shape. Thus it seemed appropriate to conduct a detailed quantitative and statistically verified experimental evaluation of distinct environmental impacts which may influence the amoeba shell.
Expriments with Trinema lineare and Cyclopyxis kahli. A two-fanctional laboratory experiment with clonal cultures of Cyclopyxis kahli and Trinema lineare which combined food supply (exact doses of yeast or bacteria) and temperature (constant 15 or 20 degrees C) crosswise revealed numerous effects of shell and aperture size as well as interactions between food and temperature (Wanner 1994; Wanner and Meisterfeld 1994; Wanner et al. 1994; Foissner and Wanner 1995). Complex experimental sets demonstrate that even a small increase in temperature causes a significant shell size reduction, which is highly reversible and reproducible (Wanner and Meisterfeld 1994; Wanner 1995). A complex of set of consecutive experiments with Cyclopyxis kahli (about half a million shells were investigated) allowed a detailed statistical analysis of interactions between and adaptations to several environmental factors. Significant changes in culture growth (lag time, generation time, final culture density) and shell size parameters were primarily dependent on food supply and temprature, whereas insecticides had a minor but still significant influence. Lime and mineral fertilizer had no direct effects, but were acting indirectly by promoting food density. Interactions between all tested factors occurred frequently, but no consistent adaptation or selection phenomena could be observed. The shell size effects were reversible within a few days by changing environmental conditions (Wanner 1994; Wanner et al. 1994; Foissner and Korganova 1997; Foissner and Wanner 1995). Because testate amoebae revealed a marked resistance to environmental stress factors, there may be no need to evolve particular adaptation mechanisms. Further investigations are essential to clarify this. In this context, the infuences of illumination on shell size properties was also investigated. Cultures of Cyclopyxis kahli, either kept in darkness or illuminated, did not show significant differences in shell size, but cultures additionally treated with a pesticide demonstrated significantly smaller shells than the respective untreated or dark cultures (Wanner 1994). Because no selection of smaller (here a result of higher temperature) or larger shells was observed in the respective subcultures, shell size modifications are assumed. A possible explanation for the observed smaller amoebae shells with proportionally larger pseudostomes caused by increased temperature may be the temperature-dependent characteristics of diffusion and respiration. An increase of approx. 10 degrees C doubles cellular respiration, while oxygen diffusion within the cell increases only proportinally to the absolute temperature (Fenchel 1990). Testate amoebae may respond to an increase in temperature by extending their active respiratoric surface in proportion to the cell volume, resulting in smaller shells with larger openings. To conclude, all the experimentally altered environmental factors did significantly affect shell size (of the respective daughter cells) - but not morphology - in a reversible and definite way, either directly or indirectly.
Variation in cell size. Variation in mean cell size in protist populations was examined as early as 1908 by Popoff, who noted an inverse relationship between temperature and cell/macronucleus size in the ciliate Frontonia leucas, and Harding (1937), who studied food-dependent cell size alterations in the ciliate Glaucoma sp. Environmentally induced cell size alterations had been also observed by Choi and Peters (1992). Cold-water isolated of the heterotrophic nanoflagellate Paraphysomonas imperforata had the largest volumes at the coldest growth temperature. Because of the characteristics of flow cytometry used in this study, it was possible to detect even small (but reproducible and statistically significant) changes in cell size (e.g. 8.2 um at -1.5 degrees C, 8.07 um at 6 degrees C, and 7.89 um at 15 degrees C). In contrast, Baldock et al. (1980), who kept six species of naked amoebae at four different temperatures, found no consistent relationship between cell volume and temperature. This applies also to the planktonic foraminifer Orbulina universa, which was cultured at four different temperatures between 19.5 to 28 degrees C, the range at which this species is abundant in the ocean. Although average final shell length (594-669 um) and shell morphology (increased frequently of individuals forming a second spherical chamber) were significantly affected, largest shell sizes were observed for the two intermediate temperatures. This may be due to other factors more important than temperature (Caron et al. 1987). Furthermore, species-dependent optimum growth temperature and culture conditions have to be taken into consideration (e.g. James and Read 1957).
The reversible nature of variability. The reversible nature of variability in protists was demonstrated by Heal (1963), who induced an increase in shell size in the testate amoeba Nebela tincta after rewetting its habitat (Sphagnum mosses). Although he did not speak of different generations, he must have sampled consecutive generations of testate amoebae, because his experiment went on for 91 days. Thus he observed shell size alterations that referred to the respective daughter cells. Schonborn and Peschke (1988) and Schonborn (1990) showed that the short-spined, soil dwelling testate amoebae Euglypha ciliata is able to redevelop long spines when kept in aqueous soil cultures. Within 48 days, the mean spine length of the daughter cells increased from 5.12 to 12.8 um.
Further environmental influences. Besides food, temperature and various drugs or pesticides, particle size in lake sediments (Schonborn 1968) affects size distribution of the amoebae. Berger et al. (1985) demonstrated in a field experiment that Trinema complanatum was significant smaller in size in compacted soil. Finally, the deficiency of silicate (Hegner 1919; Netzel and Grunewald 1977; Anderson 1990), calcite (Casper and Schonborn 1985), or the quality/lack of shell building material for agglutinated forms (e.g. Ogden and Meisterfeld 1989) in the surrounding medium causes striking alterations or deformations in the shell structure. To summarize, interacting environmental factors either natural or "anthropogenic", affect especially shell size of (the daughter cells of) testate amoebae, acting directly or indirectly in a reversible mode.
Taxonomic implications
There are qualitative shell characters useful for distinguishing taxa, as discussed in Ogden and Meisterfeld (1989). Besides biometrical data, surface composition (in combination of knowledge of the general biology and ecology), shell matrix properties, apertural structure, and cytoplasmic features as size and structure of the nucleus, number and location of the nucleolus, are suitable for species differentiation. As shown above, evironmental factors can especially affect shell size and aperture, which are relevant for an exact definition of closely related taxa e.g. ecologic and geographic races (see Wanner 1994; Bobrov et al. 1955; Foissner and Korganova 1995). Thus, identification and assessment of specific environmental factors responsible for the shell variability of distinct species or races are necessary prerequisites for further taxonomical and ecological work on testate amoebae.
Examples for variation in testate amoebae. Already in 1864, Wallich observed that shell size in Difflugia spp. is "modified to an extraordinary degree by the nature of the localities in which they happen to be found" with respect to direct influences via food supply, and climatic conditions which may act indirectly by changing sustenance. Penard (1902) mentioned polymorphism in Centropyxis constricta and Trinema spp. Leidy's minute field investigations in 1879 caused him to critisize his famous colleague Ehrenberg mentioning (p. 227) that "Trinema enchelys is very variable in form and size, and thus helped Ehrenberg to swell the long list of his Difflugias and Arcellas". Leidy himself accepted only one species, Trinema enchelys, within he whole genus Trinema because he observed numerous transitions between different morphos. Finally, Penard (1890) clearly defined Trinema enchelys. T. lineare, and T. complanatum (discussed in Penard 1902 and Cash et al. 1915). Up to now, numerous authors draw attention to the fact that many "species" of the testate amoebae may not represent "true" species but different morphos or variations of the same taxon. Bobrov et al. (1995) conducted a detailed morphological comparison of shell size characteristics of two geographically isolated populations of Nebela militaris, Hyalosphenia papilio, and Trigonopyxis arcula sensu lato. Only the first two taxa revealed statistically significant differences between the Siberian and Ontario populations, while T. arcula sensu lato (including T. arcula major and T. minuta) may be better considered as a single polymorphic complex rather than different species. Generally, the authors stressed that shell morphology measurements alone are not sufficient for delineating species. Additional morphological, ecological and genetic studies should be undertaken on the same material to improve taxon-specific differentiating.
Variation and synonymy. Cowling (1994) reviewed the relationship between body size/shape of soil protozoa and environmental influnces, and presented data about intraspecies variation and adaptations. Decloitre (1964) presented an impressingly long list of aberrant and variable forms within numerous genera and species (e.g. Nebela, Difflugia, Centropyxis, Cyclopyxis, Cyphoderia, Arcella, Assulina, Trinema, and Euglypha). Consequently he recommended to be careful with hasty descriptions of new taxa based on aberrant forms. Using morphometry, light microscopy and SEM, Foissner and Korganova (1995) redescribed the shell morphologies of terrestrial testate amoebae. They concluded that morphometic criteria are of very limited taxonomic value, and only applicable for species separation if they are very distinct. Generally, to avoid an oversplitting into apparently closely related species, they suggested that individuals of field populations very likely belong to the same species if the coefficient of variation of the main metric shell characters (e.g. shell and pseudostome/aperture size) does not exceed 30-50%. Additionally, they recommended at least one extra reliable morphological character for separation. Thus Cyclopyxis kahli cyclostoma, C. kahli obliqua, C. dispar, C. dulcis, C. insolitus and C. profundistoma is synonymized with C. kahli and Cyclopyxis gigantea, C. bathystoma and C. kahli grandis synonymized with C. intermedia. Supplementary discussions are given in Luftenegger at al. (1988), Wanner (1988), and Luftenegger and Foissner (1991), who characterized numerous soil testate amoebae morphometrically. Heal (1961) mentioned difficulties in separation of closely related species, e.g. in Centropyxis spp., Nebela spp., Heleopera spp., and Phryganella spp., and recommended additional culture experiments to distinguish species. In 1963 he stressed that, although morphometric studies have been made to assess the validity of various species, it is necessary to obtain more information about the reasons of variation. In this work, he measured about 1060 tests of the Nebela tincta-bohemica-collaris group from Sphagnum and found that Nebela tincta and N. flabellulum are distinct species, but N. collaris sensu lato includes N. bohemica. Remarkably, he demonstrated that the size of Nebela increased when wet Sphagnum was flooded exprimentally. Thus it was concluded that Nebela tincta in the range of 75-95 um is distinct from larger forms, but specimens ranging from 95-155 um in length form a cline due to the variation in water content of the habitat. Luftenegger et al. (1988) also studies morphometric variation in the Nebela tincta-parvula-bohemica-collaris group and separated this species complex on the basis of shell size into N. tincta (with lateral pores)/N. parvula (no lateral pores, both taxa about 80-110 um) and N. bohemica/N. collaris (about 100-200 um). However, some intermediate forms occurred in the populations, too. The presence of absence of lateral pores as a valid taxonomical criterion and further taxonomical problems in Nebela are (controversially) discussed in e.g. Leidy (1879), Cash and Hopkinson (1909), Deflandre (1936), MacKinlay (1936), Hoggenraad and de Groot (1937), and Heal (1963). But -as stated in Luftenegger et al. (1988)- as long as no better criteria are known, it may be useful for separation. With regard to the change in size within a species, Jung (1936) stated after measuring 21 shells of Assulina muscorum, that this species becomes smaller with decreasing water content of the habitat. A quite controversiall discussed case study of intraclonal variability in testate amoebae was presented by Medioli et al. (1987). They claimed that intraclonal variability represents the "minimum" variability that can be expected in a given (uniparental) species. Only morphotypes falling outside the interval of variability will belong to other species. Based on this hypothesis, they synonymized about 100 taxa to the single species Difflugia tricuspis. Unfortunately, as discussed in Ogden and Ellison (1988) and Ogden and Meisterfeld (1989), they did not consider additional data, as those belonging to the idiosome bearing genus Netzelia. Extensive information about morphometry, shape and structure of testacean shells is given by Odgen (1980, 1981, 1983, 1984, 1999), Ogden and Hedley (1980), Ogden and Couteaux (1987), Ogden and Meisterfeld (1989), and Schonborn (1992). Generally, it was underlined that the combination of detailed species descriptions and information deriving from clonal cultures concerning the natural variability of shell formation may be helpful in finding reliable specific features. In turn, the number of new species based on poor diagnoses should be restricted.
Variation in different taxa and shell characteristics. To sum up, description (or redescription) of species as well as species-based ecological work is only useful if reliable biometric data are provided. In this context, the analysis of the coefficients of variation is of special interest, because it allows direct comparisons and evalutions of differently sized shell characteristics. However, it must be taken into consideration that different taxa as well as different shell characteristics within a given taxon do vary divergently. For example, the shell morphology in Centropyxis spp. is more variable than in Arcella spp., and in both genera the aperture ("pseudostome") is more variable than the shell size (Heal 1963; Meisterfeld 1978; Luftenegger et al. 1988; Wanner 1988, 1991, 1995; Wanner and Meisterfeld 1994). Univariate as well as multivariate statistics of main shell characteristics of terrestrial testate amoebae showed that the smallest cubic axis (the thickness or height) of the shell varied independently as compared to other axes (Wanner 1988, 1991). This indicates an adaptation of the amoeba shell to the thin water-film covering the soil substrate (e.g., Heal 1963; Bonnet 1964; Schonborn 1966; Foissner 1987; Wanner 1988). The feature complex "pseudostome" also shows a strong variablity. The respective diameters and the invagination are not coherent because they vary independently (Wanner 1988, 1991). Furthermore, a computerized image-analysis of the shell of six populations of Schoenbornia humicola revealed no significant differences in shell size, while shell platelets showed highly significant differences in form and size (6061 platelets of 110 shells had been measured), indicating habitat specific influences (Wanner 1991). Moreover, the platelets may also be derived from other species (Schonborn et al. 1987). Finally, it had been proved quantitatively using image-analyzed clonal cultures of testate amoebae (Wanner 1994, 1995; Wanner and Meisterfeld 1994; Wanner et al. 1994), that all tested taxa, clones and cultures showed a taxon-specific, highly reproducible and reversible variability influenced by interacting culture conditions. However, it is important to know that, in spite of the "extreme" experimental conditions in culturing Cyclopyxis kahli, neither pronounced alterations in shell morphology nor an exceeding of the shell size range of the respective "wild" taxon could be observed. The latter applies to all clones and taxa investigated (Tabel 1).
Species delimitation and reasons for variation in testate amoebae. Based on the above collected data, the following questions arise: Which are the feature charaterizing a species, and in which way are these features affected by environmental factors? On one hand, habitat structures may stabilize the morphological characteristics of a taxon, on the other hand micro-scaled variable environmental factors may infleunce the morphological characteristics in various ways (Schonborn 1989, 1992). The current biological species-concept may be insufficient for organisms with a predominant asexual reproduction (Meisterfeld 1979; Tibayrenc et al. 1991; Poljansky 1992). Alternatives, like the phylogenetic species concept (Cracraft 1983), or the evolutionary species concept (Simpson 1961), have some practical disadvantages for parthenogenetic species and asexual clones, as discussed in Quicke (1993). Additionally, with the evolutionary species concept, it is not possible at any one time to know the historical fate of an extant lineage, and phylogenetic species concept may result in an extreme oversplitting of species, as compared to the more traditional biological species concept. Working with an "ecological species" - a morphological divergence is interpreted as an expression of a reproduction community - this problem may be circumvented to a certain degree (Willmann 1985). To conclude, only a more or less temporary solution based on a combined approach, including criteria for monophyletism, ecological niches and morphological and genetical similarities, will be successfull (Zakrys 1997). Obsevations on clonal cultures of testate amoebae indicate high variations in shell characteristics which are used for species determination (discussed in e.g., Heal 1963; Meisterfeld 1979; Schonborn and Peschke 1990; Wanner 1994; Foissner and Korganova 1995), problems which also occur in other protist groups (e.g., Zakrys 1997). However, there are existing stable characteristics useful for species discrimination, but these features have to be estimated in each individual case (Meisterfeld 1984, Ogden and Meisterfeld 1989). Yet, due to missing culture and molecular data, the actual reasons for variation in testate amoebae can be proved in a few cases only. To summarize, variable shell features of testate amoebae ae commonly used for taxonomic purposes. As discussed above it is likely, that a considerable part of the present known taxa (about 1600 species) may be only variations of "true" species (controversially discussed in Medioli et al. (1987), Ogden and Ellison (1988), Ogden and Meisterfeld (1989), Foissner and Korganova (1995). Assuming this hypothesis, there is need to know in which way species may split in different morphs or variations. (a) The "classic" studies on clonal cultures postulated either persistent modifications ("dauermodifications" - Jollos 1917, 1924) or mutations (Jennings 1916; Root 1918; Hegner 1919, 1920; Reynolds 1923) as a basis for variation. (b) Chardez and Leclercq (1963) separated Euglypha strigosa into four habitat-specific ecotypes with different spines: Euglphya strigosa heterospina (aquatic), Euglypha strigosa strigosa (Sphagnum), Euglypha strigosa muscorm (mosses), and Euglypha strigosa glabra (terrestrial). Schonborn (1983) interpreted these morphs as "Dauermodifikationen", because in his aqueous cultures, spineless terrestrial shells transformed into spiny morphs. Putative extra-chromosomal factors firstly substantiated the concept of "dauer-modification", but Danielli (1959) showed that mainly chromosomal genes are responsible for most features of amoebae. Thus persistent modifications occur, but are likely to paly only a minor role (Schonborn 1983, 1992). (c) Subsequently Schonborn developed the concept of an "adaptive polymorphism" as explanation for culture experiments on stability and selection in varieties and morphs of testate amoebae, namely Trinema complanatum and Euglypha laevis. Four different morphs of Trinema complanatum isolated from spruce forest humus and soil samples (small-rounded, large-rounded, waisted, and small morph), indicated a discontinuous size polymorphism. After Schonborn (1992) adaptive forms are hidden behind this discontinuity. In clonal cultures of Trinema and Euglypha, the features of the morphs remained relatively constant, but over the course of several months other morphs occurred. These morphs could be transformed into each other by selection, while "pure" shell size variability, as shown for Trinema complanatum, may have a modificatoric basis. Furthermore, in clonal cultures of Euglypha with round pseudostomes ("E. rotunda") individuals also occurred with oval pseudostomes ("E. laevis"). Thus Schonborn (1992) synonymized the later described E. rotunda (for practical reasons, E. rotunda was maintained in Table 1). However, only an estimation of the genotypical conditions will allow futher knowledge of variation, as discussed for Euglypha spp. (Zakrys 1997) and Amoeba proteus (Poljansky 1992). Besides molecular approaches, which are essential for the future, clonal cultures of testate amoebae (informing about intraclonal variability) are important for taxonomy too (e.g. Ogden and Couteaux 1988). Furthermore, ultrastructural research on shell structure, cytoplasm (pseudopodia, nucleus), and organic matrix are necessary (e.g. Stout and Walker 1976; Netzel 1977, 1983; Ogden 1979, 1981, 1983, 1988, 1991; Ogden and Couteaux 1987; Ogden and Meisterfeld 1989, 1991; Ogden and Pitta 1989).
Conclusions
"Conventional" bioindication with testate amoebae - using community structure parameters like abundance, dominance, species spectra, and biomass - is successfully established since a long period. As compared to naked amoebae, flagellates, and ciliates, no costly or time-consuming methodology is necessary, since direct observation of an aequeous suspension is sufficient for qualitative and quantitative analysis. With respect to forest soils or agroecosystems, their relatively small species richness, as compared to other protist groups, faciliates practical work on bioindication. However, field investigations based on shell size alterations may not be practicable at the moment, since even few environmental factors produce complex morphometric alterations. In contrast, laboratory tests using clonal cultures of testate amoebae seem to be more promising. Experimentally changed evironmental conditions resulted in distinct alterations in aperture and shell size, which were shown to be highly reproducible and reversible within a few generations. If reliable qualitative shell characteristics are available, shell size variability may cause no severe taxonomic problems, but regarding closely related taxa, where separation occurs mainly by size characteristics, definition of species and bioindication may be impeded. To sum up, intraclonal variability plays an pivotal role in basic and applied research. On one hand, it implies some current taxonomic and ecologic problems, but on the other hand, it offers new and promising possibilities for the future. Based on the above referred data, the following research objective are proposed:
- the search for additional qualitative species characteristics, morphological as well as molecular features, as a basis for species delimitation;
- the comprehensive evaluation of clonal variability in numerous taxa of testate amoebae to provide basic information for taxonomic and ecologic evaluations;
- the development of routine laboratory (toxicity) tests by means of shell morphometory alterations as a new tool for indication - e.g. to replace/supplement animal experiments;
- the establishment of molecular methods for the characterization and delineation of (contaminated) field populations by amplifying a sequencing DNA deriving from single amoeba nuclei;
- the establishment of contamination-free cultures to detect sexual processes in testate amoebae using RAPD-derived DNA-fragment patterns. Such cultures may be good candidates for a subsequent ultrastructural search for meiosis, and may help to clarify the above discussed problem of species definition;
- finally, basic research on so-called "dauermodification', seemingly forgotten by most scientists, may be renewed using testate amoebae as convenient model organisms. (ref. ID; 2852)
Dr. Anatoly A. Bobrov Address: Department of Soil Science, Moscow State University, Moscow, Russia
ref. ID; 4863
Dr. Colin G. Ogden (the deceased) ref. ID; 890, 891, 2022, 2024, 2290, 2291, 2292, 2293, 4727, 4799, 7506, 7610, 7701, 7745
Dr. Didier Chardez Address: Attache de Recherche, A la faculte des Sciences Agronomiques-Gembloux (Lab. Zoologie generale et Faunistique) 3, Chaussee Romaine, 4252 OMAL, Belgique
ref. ID; 2122, 2392, 2818, 7505
Dr. Edward A.D. Mitchell Address: Laboratory of Soil Biology, University of Neuchatel, Rue Emile Argand 11, CH-2009 Neuchatel, Switzerland
ref. ID; 6728, 6790, 6793, 6795
Dr. Hada Yohine (1905-1993) ref. ID; 3496, 5026
Dr. Jim Green Address: Zoology Department, Westfield College, Hampstead, London, England
ref. ID; 2356, 2683
Dr. John McN. Sieburth (retired) ref. ID; 3626
Dr. Kenneth H. Nicholls Address: S-15 Concession 1, PR #1, Sunderland, Ontario, Canada LOC 1HO
ref. ID; 4910
Dr. Louis Beyens Address: University of Antwerpen, Campus Drie Eiken, Department Biology, Polar Ecology, Limnology and Paleobiology, Universiteitsplein 1, BE-2510 Wilrijk, Belgium
ref. ID; 2122, 2392, 2818, 4738, 4763, 7505
Dr. Manfred Wanner Address: State Museum of Natural History, POB 300154, D-02806 Goerlitz, Germany
ref. ID; 2030, 2852
Dr. O. Roger Anderson Address: Biological Oceanography, Lamont-Doherty Geological Observatory of Columbia University, Palisades, New York 10964
ref. ID; 4245, 7360
Dr. Paraskevi Pitta Address: We do not have the address.
ref. ID; 891
Dr. Paul E. Hargraves Address: Graduate School of Oceanography University of Rhode Island, Narragansett, Rhode Island 02882-1197, USA
ref. ID; 3626
Dr. Paul Van Oye (1886-1969) ref. ID; 2230, 2233
Dr. R. Neal Band Address: Department of Zoology Michigan State University, East Lansing, Michigan 48824, USA
ref. ID; 3680
Dr. Segei O. Skarlato Address: Head of the Laboratory of Cytology of Unicellular Organisms Institute of Cytology RAS Tikhoretsky Ave., 4, 194064 St. Petersburg, Russia
ref. ID; 2031
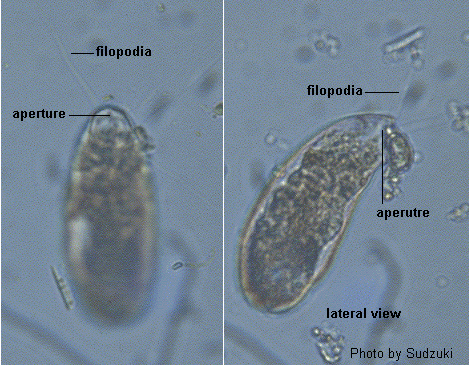
Dr. Sudzuki Minoru Address: Biological Laboratory Nippon University, Higashi-arai, Omiya-shi Saitama-ken 330-0825, Japan
ref. ID; 3057, 3536
Dr. V. Cepak Address: Czechoslovak Academy of Sciences, Institute of Botany, Section of Plant Ecology, Treborn, Czechoslovakia
ref. ID; 4823
Dr. Vassil Golemansky Address: Institute of Zoology 1 Tsar Osvoboditel blud, 1000 Sofia, Bulgaria
ref. ID; 2031, 2292
Dr. Vera Opravilova Address: Department of Biology of Animals and Man, Faculty of Science, J. E. Purkyne University, Kotlarska 2, 61137 Brno, Czechoslovakia
ref. ID; 2555
Dr. Vladimir Sladecek Address: Institute of Chemical Technology Department of Water Technology and Environmental Engineering, Technicka 5, 166 28 Praha 6, Czech Republic
ref. ID; 3343
Dr. Whilhelm Foissner Address: Universitat Salzburg, Institut Zoologie, HellbrunnerStrasse 34, A-5020 Salzburg, Austria
ref. ID; 2030, 2034, 4755
Dr. Wolfgang Petz Address: Universitat Salzburg, Institut Zoologie, HellbrunnerStrasse 34, A-5020 Salzburg, Austria
ref. ID; 2030, 4755
We cannot still accept permission to quote some description from following articles. Please allow us to reproduce. (June 30, 2018)
ref. ID; 654, 2037, 2091, 2241, 3457, 3761, 5772, 7130, 7582, 7609