No segments on the body; papillar projections; A pair of large, thick toes; caudal elevation; sensitive side-papilla; pedious glands; hairless rotatory apparatous; there is one sensitive papilla on the head. (ref. ID; 2650)
ref. ID; 3535
The general shape of the animal is moderately elongated fusiform just like that of usual notommatid rotifers of the genera, Taphrocampa, Notommata (tithasa and ometata), Resticula, Pleurotrocha (hyalina), and four allied genera, Proales (brevipes), Cochleare, Lindia (parrotti) and Proalides. Both extremities, i.e. head and tail, are usually truncated when contracted, but actually the tail ends in a degenerated foot provided with two large, protrusile toes. Since the body is devoid of lorica and transparent, the internal structures of the animal are observed with ease from outside. But in some cases the surface of the body is partly or wholly covered with dust or debris of the soil particles, and then the internal organization is difficult to observe from outside. The most particulate characters of the present genus are: first, the papillar projections associated with the peculiar dust-coating habit; second, a prominent dorsal antenna and a peculiar caudal elevation; third, remarkably large-sized two toes. Besides these characters, the vestigeal foot, the type of egg-carrying, the form of mastax, etc. are also highly peculiar. These will be described in detail in the following lines. (ref. ID; 3535)
Papillar Projections: The present rotifer is, in the case of f. vermiformis, provided with several (15-30) remarkable projections on the epidermis; these are arranged in three longitudinal rows on the dorsal and in two rows on the ventral integument. Each dorsal row usually consists of about 3-7 projections, each ventral row of 2-4 of them. And, these projections as well as the body surface are highly viscid in nature. Such projections as seen in the form vermiformis have never been reported before in the literature on the order Monogononta. Things like these, however, are found in two species, namely, Tylotrocha monopus (Jennings, 1894) under the family Notommatidae and Proalides subtilis (Rodewald, 1940) under the family Brachionidae. In the former species, the processes are described as "knob-like". But, these processes are only two (one pair) in number and exclusively located on the lateral sides of the middle part of the trunk (Harring & Myers 1926, p.516). In Proalides subtilis, on the other hand, projections -"buckelartige Fortsatze" (Rodewald 1940, p.276)- are reported to be 3-5 in number and develop merely on the dorsal side arranged in a longitudinal row. Moreover, in those two species the projections are as the body surface never papillar in nature, consequently bearing no dust. It is highly probable that these species have no organs which might serve as sticky glands. Thus, not only in the number, distribution and the structure but also in the function of the organs, the present species is wholly different from T. monopus and P. subtilis. As to the dust-coating habit, it has already been reported for four species, i.e. Proalides tentaculatus, Taphrocampa selenura, Paradicranophorus hudsoni and Lindia parrotti, that the dorsal surface of the animal is highly viscous and usually covered with particles of certain materials just as in the present genus; so the body is covered with "debris divers" in Proalides (de Beauchamp, 1907), "floccose material" in Taphrocampa (Harring & Myers, 1926), "Detritus und Schlammteilchen" in Paradicranophorous (Wulfert, 1936) and "flocculent matter" in Lindia parrotti (Russell, 1947). However, in these species the marked papillar organs as those of the present species are wholly lacking on any part of their integument. (ref. ID; 3535)
Dorsal Antenna: The genus Liliferotrocha has a fairly prominent dorsal antenna with a tuft of sensory cilia at the occipital region of the animal; its shape resembles that of only three species, namely, Proalides tentaculatus, Proalinopsis caudatus and Lindia parrotti, and is different from that of all other usual notommatid genera, e.g. Notommata, Taphrocampa, Pleurotrocha, Resticula, Tylotrocha and Proales. Here, it may be worthy to mention that other Proalides-spp. and usual Lindia-spp. have no such prominent antenna. (ref. ID; 3535)
Caudal Elevation: In Liliferotrocha, there is a single, moderately large, round, lobe-like elevation on the dorsal side of the caudal region. Similar elevations have been reported in a few species, for example, Notommata (falcinella, tripus), Proales and bdelloid rotifers. However, in the structural respect, the caudal elevation of the present genus is slightly related to the so-called spur. Rodewald (1940, p.276) mentions in his species subtilis a "vorstulpbarer Fussanhang". It is highly probable that this projection on the foot is not identical with the author's elevation. Perhaps Rodewald simply mentions the toes which are very introversible. (ref. ID; 3535)
Toes: The present genus has remarkably large, sickle-shaped toes; they are as long as or a little less than one-fourth of the entire length of the body. Their size, shape and number are somewhat similar to those of Notommata (tithasa), Aspelta, Tetrasiphon, Sphyrias, Rousseletia, Euteroplea and Proales (minima), and fairly different from those of other notommatids, e.g. Taphrocampa, Pleurotrocha, Thylotrocha, Resticula and allied genera Lindia and Proalides. The present genus has no such claws as found in Brachionus diversicornis, etc. (ref. ID; 3535)
Foot: In the present genus there is no distinct boundary between the foot and the body proper, which fact does not agree with the criteria of the subfamilies Proalinae, Tetrasiphoninae, Birgeinae, Dicranophorinae and the majority of the genera under the subfamily Notommatinae. (ref. ID; 3535)
Egg-Carrying:
Quote from ref. ID; 3535
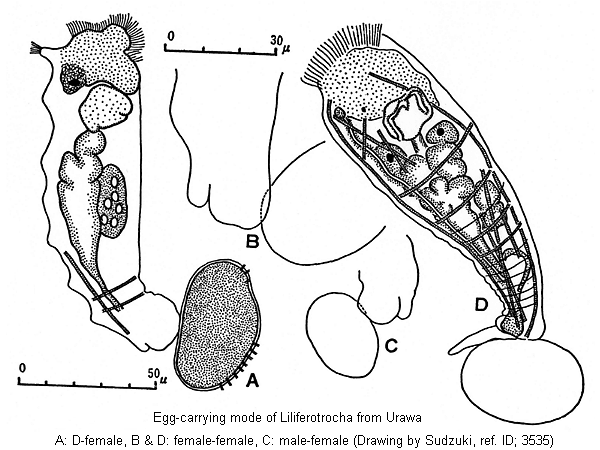
In Liliferotrocha all sorts of the produced eggs, namely, amictic, unfertilized mictic and fertilized mictic are carried attached to the caudal extremity of the animal by means of the sticky substance secreted from the style glands at the coxal part of the toes. The egg carried in this way is usually one in number even in the unfertilized mictic female. This type of egg-carrying has been observed only in two rarest species, Proalides tentaculatus and Lindia parrotti (De Beauchamp 1907, p.149, Fig.A; Wulfert 1941, p.172, Fig.B; and Russell 1947, photo.), but not found in the usual families Notommatidae and Brachionidae. In Proalides tentaculatus and Lindia parrotti, however, there are no style glands but foot glands at the coxal part of the body. On this matter, de Beauchamp assumes that in Proalides tentaculatus the foot glands play a role in the egg-carrying. (ref. ID; 3535)
Mastax: According to Harring-Myers (1926, p.416), the types of the trophi are the most important taxonomic character of the notommatid group, these authors divided the family Notommatidae into six subfamilies by the degree of departure from the original malleate type; 1) "Malleate"-like type occurring in Proalinae, 2) "Virgate" type in Notommatinae, 3) Type with an unusually developed epipharynx in Tetrasiphoninae, 4) "Cardate" type in Lindiinae, 5) Virtual atrophi type with a pair of "pseudo-unci" in Birgeinae, and 6) "Forcipate" type in Dicranophorinae. Almost similar classifications founded chiefly upon the trophi types have been accepted by Remane (1933), Pennak (1953) Donner (1956) and Voigt (1956-57) for the taxonomy of this group of rotifers into families rather than subfamilies. The trophi type of Liliferotrocha appears to be changeable with stages of development; the embryonal trophi are of the malleate type, while those of the adult are subject to great modifications as in the uncinate type of the mastax. In the young, the fulcrum is extremely short, ending in an expanded plate or basal apophysis like that in Proalinae and also Lindia parrotti; the rami are triangular in shape and without alurae such as of tentaculatus (Wulfert, 1941); the unci have only a few teeth in the adult, while, in the embryo, they have about 7-8 teeth like those in the adult of Proalides and Proales. The manubria are both long, bent inwards symmetrically, provided with a spatulate expansions; and, certain spine-like projections as found in Pleurotrocha hyalina (Wulfert, 1939) are observed at both shoulders. The epipharynx which is found in Lindiinae and Tetrasiphoninae is not recognizable in this genus. From the features of the trophi just mentioned above, the present genus seems to be hardly related to almost any of the notommatid subfamilies, e.g. Dicranophorinae, Notommatinae, Birgeinae, Lindiinae, Tetrasiponinae (Lecanidae), except only Proalinae, especially Proales. The last mentioned subfamily is now treated under the family Lecanidae for the reason that it has the malleate type of the mastax. (ref. ID; 3535)
Neck Segment: In the majority of the notommatid genera, there are found some remarkable constrictions which separate the head or the neck from the abdomen in front of, or behind, the mastax. The present genus is, however, totally lacking this organ as Lindia parrotti and Proalides. In this point, Liliferotrocha lies very remote from all the notommatid subfamilies except Tetrasiphoninae and one genus Taphrocampa under Notommatinae. (ref. ID; 3535)
Auricles: In the central group (cf. Harring-Myers) of the Notommatinae, namely, Notommata, Taphrocampa, Tylotrocha, Itura and also Lindia (s. str), this organ is reported to be well developed. In the present genus, however, any trace of it is not observed. (ref. ID; 3535)
Retrocerebral Organ: In the present genus, neither retrocerebral sac nor sub-cerebral glands are found as in Birgeinae; otherwise, they are markedly developed in many genera under the subfamily Notommatinae (except Pleurotrocha and Wulfertia) and Dicranophorinae as well as in all the genera of Lindiinae and Tetrasiphoninae. (ref. ID; 3535)
Other minute character: These is a fine rostrum-like projection at the tip of the head. The palp organ which has been reported in Rousseletia is not found. The corona is an oblique disc with well developed marginal cilia. The buccal field is comparatively small and evenly ciliated. The mouth is ventrally situated at the usual site, and on each side there are found certain projected regions, which may not be observed from the dorsal side. The post-oral portion does not project as chin, contrary to the genera Notommata, Lindia (except, parrotti) and Eothinia. The brain is situated at the normal site of the head, at the posterior end of which is observed one cervical eye spot, which appears to have a lens. In this point, Liliferotrocha differs from Tylotrocha, Drilophaga, Sphyrias, Eosphora and the majority of Dicranophorinae. The oesophagus is slender (15-23 µm) and ciliated as in Taphrocampa, Notommata and Proales (?), and unlike Proalides, Proalinopsis, Pleurotrocha, etc. No remarkable salivery glands are recognized in Liliferotrocha, while they are found in Drilophaga, Resticula, Sphyrias, Eosphora, Euteroplea, Paradicranophorus, and also in Lindia, Notommata, Taphrocampa. The gastric glands are present at the normal position and of normal spherical shape, usually containing 6 nuclei each, and open into the stomach through a short duct, 5 µm long and 1-2 µm diameter. On this structural point, the present genus is different from such genera as Birgea, Pleurotrocha, Tylotrocha, Itura, Euteroplea, Wierzejskiella, Paradicranophorus, Proales, Cephalodella and Resticula. The intestine and the stomach are well distinguished. This is not the case in the subfamily Proalinae and also in such notommatid genera as Pleurotrocha, Rousseletia, Baltro, Lindia (parrotti), etc. The lateral antennae, situated at the posterior level of the first third of the body, are usually spindle-shaped, each provided with some sensory hairs. In this point, too, Liliferotrocha is not like Drilophaga, Taphrocampa, Rousseletia, Resticula, Euteroplea, Eothinia, Proalides and also the subfamilies Tetrasiphoninae and Lindiinae. The protonephridia are recognized as two lateral canals with a few flame bulbs and opening into a large bladder at a level between about 1/5 and 1/7 of the body from the posterior end, and the excreta are discharged into the cloaca through a short duct. Proalides has been reported to have neither protonephridia nor flame bulbs (de Beauchamp). The cloacal opening is dorsally located at the base of the toes. The vitellarium is large, having usually 8 nuclei in it. It is situated at the right side of the trunk and occupies most part of the body cavity, extending from above the stomach to below the intestine. The general contour of the dormant egg, which is considered to be an important generic character of the rotifers, does not seem to resemble that of Proales-spp. Further, the three kinds of eggs of the present genus can not be compared with those of other notommatid rotifers, since the corresponding eggs of other genera have not been described nor figured to date. (ref. ID; 3535)
Notes
Liliferotrocha is sharply distinguished from P. tentaculatus in having 1) caudal elevations, 2) particular papillar projections, 3) well developed toes, 4) lateral antennae, 5) a protonepharidial system, 6) a particular musculer system, and in wholly wanting 7) annulations and 8) the peculiar pattern on 5th annulation on the trunk. Further, it shows peculiarities in the structure of such digestive organs as 9) oesophagus (without cilia in tentaculatus), 10) gastric glands (with large-size nuclei in tentaculatus) and 11) ramus and fulcrum of the mastax (mallleo-ramete in tentaculatus after de Beauchamp). Of these, 3), 4), 5), 9) and 10) seem to have been overlooked by previous researchers using only a few or fixed individuals, but 1), 2), 6), 7), 8) and 11) are of great importance and well deserve the generic distinction. In this connection, it may be worthy to mention that there exist some morphological differences between the specimens identified as tentaculatus by de Beauchamp (1907) and those by Wulfert (1941). Such differences are found in the number of annulations, the presence or absence of chin, position of the eye, structure of fulcrum with apophysis, rami with alurae, number of vitellarium nuclei, situation of cloacal pore, and so on. Probably, Wulfert's specimen is distinct on the specific level from tentaculatus s. str. Liliferotrocha is distinguishable also from another proalid species, subtilis, in the characters of 1) caudal elevation (not toes), 2) rami of mastax, 3) papillar projections (developed also on the ventral side of the integument), 4) prominent dorsal antenna, 5) dust-coating habit, 6) type of bladder, 7) shape of toes, 8) foot glands (size), 9) corona (terminal in subtilis), 10) fulcrum, intramallei, and 11) ecology. Since no description is available of retrocerebral organ, gastric glands, egg-carrying type, vitellarium, protonephridium, etc. of subtilis, we cannot compare our species with subtilis on these points. The present author nevertheless quite agrees with Rodewald in assuming that his species, beyond doubt, belongs to the genus Proalides. On the other hand, genus Liliferotrocha differs clearly from Russelletia (= Lindia) parrotti in having 1) papillar projections, 2) caudal elevations, 3) lateral antennae, 4) toes of special shape and size (like L. tecusa in parrotti) and in lacking 5) the annulations 6) the semi-circular retractile chin, again also in 7) the minute structure of the digestive organ (stomach is not separated from the intestine in parrotti), 8) the type of the trophi, besides 9) some ecological characters (habitat, occurrence, etc.). (ref. ID; 3535)
Body elongated cylindrical with unclear transverse bands. Dorsal antenna large, clearly visible in lateral view; toes short, slender, pointed. Eggs carried on posterior end of body. Amictic eggs slightly smaller than resting eggs. (ref. ID; 1806)
Our specimens were characterized by a weakly developed rotatory apparatus, exempt of tufts of cilia. The dorsal sensitive papilla was large, provided with some sensory cilia at the extremity. Two nerves issuing from the cerebral ganglia and one muscle able to generate a certain retraction of the papilla penetrated far into the papilla. On both sides, the head was provided with two sensitive papillae. The eye, situated above the cerebral ganglia was either simple or double. The trophi were composed of a short fulcrum, ending in an expanded plate and two rami -square shaped and similar to spinned plates- that were irregular distributed; sometimes, these were triangular in shape. The manubria are long, ending in a widened plate with sharp external extremities. In side-view, the manubria are (thickened anteriorly) showing a aperture similar to window. The unci are thin -stick-like - distributed by 7 or 8 on each side and closely grouped together, forming a plate with deep grooves. Several strong distinct muscles are fixed on the rami. It is worth mentioning that the description of the mastax given by Sudzuki corresponds to the case when this one is treated with sodium hypoclorite or another substance that permits the breaking up of the animal into constituent parts and their displacement off the normal position. Usually, the pieces of this apparatus are arranged regularly. The digestive and excretive apparatus as well as the muscular system agree to the description made by Sudzuki. The vitellin gland is cular system agree to the description made by Sudzuki. The vitellin gland is clearly bilobated. Toes are large, slightly retractile. Usually, the first toe is hollow, the second contains an internal cytoplasmic formation which looks like the reservoir and canal of a pedious gland. Sometimes, the poster lobe may be bilobated. (ref. ID; 2650)
Comments
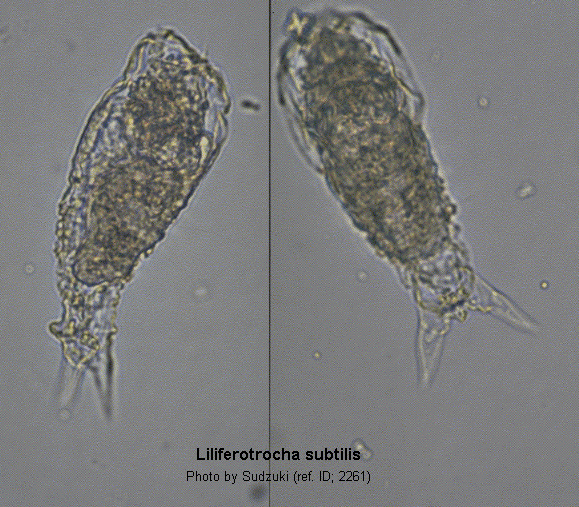
Rodewald (1940) described a new species of Proalides -Proalides subtilis- from the Siut-Ghiol Lake. M. Sudzuki (1959), Japan, gives a detailed description of Liliferotrocha urawensis n. g., n. sp., on the basis of a rich and varied material collected through a 7 years-term. Despite the fact that Sudzuki has given a more complete and detailed description of urawensis, Rodewald had the priority of its description; therefore the species name should remain Liliferotrocha subtilis (Rodewald, 1940). According to Sudzuki, the species is polymorphous, as three morphological types could be found within the same population; they have been called by Sudzuki -naturally without any taxonomic value- f. notommatoformis, f. vermiformis and f. tintinnoformis. (ref. ID; 2650)
Measurements
Total length 97-156; maximum width 30-42; maximum thickness 32-36; toes 13-30; caudal elevation 4-5; trophi 15-18/19-22; fulcrum 2.2; rami 2; manubria 5.2; unci 3.2-3.8 µm. (ref. ID; 2650)
The body is moderately elongated fusiform, tapering to the caudal extremity. The greatest width, which lies near the neck region or at the level of the first fourth of the boy length, is about one-third to one-fifth of the total length. The narrowest part is about one half to one-third of the widest part, lying nearly at the level of the last fourth or the last fifth of the body excluding the toes. The body is illoricate and the integument is soft, very flexible, with no annulations. The body is originally transparent, but among the specimens there are a lot of forms distinguishable from the original type owing to the development of the papillar projections, which are associated with the dust-coating habit. Namely, in the present species, three morpho-ecological forms are observed concerning chiefly the papillar projections: -
In one form, the animal has no nipple-like papilla projections, and bears no dust or soil particles on its whole surface. It looks as a usual notommatid rotifer. This fundamental form may be called here forma notommatoformis.
In the second form, only the papillae are covered with dust or soil particles The animal resembles a caterpillar with some strongly developed setae. This form may be named here forma vermiformis.
In the third form, the internal organization is hardly seen from outside owing to the presence of soil debris on the whole body surface. The animal while swimming somewhat gives an appearance of a tintinnoid protozoan, Tintinnodium fluvitiale (Hada, 1938). Thus, the name tintinnoformis is proposed. These trimorphic features are the most peculiar characters of the present species.
At the tip of the head or on the aciliated apical plate there is observed a tripartite rostrum-like projection. Close to this organ are found two pairs of rather long cilia. A weakly developed ciliary field is present on the ventral side of the head; the buccal field is not so large and evenly ciliated, in which a small mouth, 1 µm in diameter, is situated. The auricles are wanting. No neck segments. Therefore, head, neck and abdomen are not sharply separated from one another. The brain or cephalic ganglion, 15x8 µm in size, is situated at the central position of the head. At the posterior end of the cephalic ganglion (i.e. at about one sixth of the body length) there is found an eye spot which is partly red partly yellow (with a lens as in Testudinellidae). No traces of the retrocerebral organ. The pseudocoel is well developed at the head region just beneath the epidermis. The mastax is 23-25x13-15 µm in size. The trophi, which are bilaterally symmetrical, are in the adult so reduced that we could not decide its exact type, but in the young it is of the malleate type; the fulcrum is remarkably short, 6-8 µm length, ending in a basal, expanded plate. The rami triangular, 4-5x3 µm in size, without alurae. The unci, 5-7 µm in length, appear to have a few teeth (often 5-6) in the young. The latter organ is slender, clubbed at the tip, and tapers off towards the posterior margin. In the embryonal stage, however, the unci seem to have seven to eight teeth. The manubria are 12 µm in length, slightly curved inwards near the edge, and end in an expanded oval plate. No epipharynx is present. Dorsal antenna is a remarkably well developed organ, 11x6 µm in size, having a tuft cilia (6-10 µm long). It is situated at the normal site. From the cephalic ganglion to this organ run two pairs of nerve cords. The lateral antennae, provided with spine-like sensory hairs, are situated at the anterior limit of the last third or nearly in the middle of the body. The oesophagus is ciliated, considerably long, 15-23x3 µm in size. Around this organ there are found three nerve cords, each having 2-3 ganglia, On both sides of the distal part of the oesophagus, there is a pair of elongated gastric glands, 10-20x7-10 µm in size, consists of about 35 cells, each of which bears comparatively short cilia (5 µm in length); the stomach leads to the unlobed intestine which is represented by a single slender duct (23x15 µm), the inner epithelium of which is provided with rather long cilia, more than 12 µm in length. The rectum is cylindrical, 30 µm long, and the anus is placed at the upper base of the toes. There are two pairs of longitudinal muscles and more than four pairs of circular muscles. The protonephridial system is rather obsolate. However, two lateral canals with two pairs of flame bulbs are clearly seen immediately over the stomach. The bladder, 20-30x8-15 µm in size. Contracts irregularly about 6 times per minute, while in the embryos it contrasts rhythmically once per 10 sec, and in the very young embryos once per 90-107 sec. (usually 95 sec). This organ, together with cloaca, opens to the exterior. The large vitellarium is situated on the right side of the body, occupying the most part of the body from the gastric region to the distal part of the intestine; it is 38-50x16-17 µm in size and occupies almost one third of the entire body cavity. The organ consists of two portions, the one with 5 and other with 3 nuclei. Non of these nuclei shows any polymorphic character as found in Filinia longiseta limnetica and Asplanchna sieboldii (Sudzuki 1958). At the caudal region there are often observed some evanescent transverse folds as found in the sessile rotifers. The region corresponding to the foot is not so markedly separated from the body, and usually no distinct segments are recognizable in it. The toes are pretty large, 17-20 µm long and 8-12 µm wide. They are retractile and may be completely withdrawn into the body for some hours, but, when protruded, each toe displays always a sickle-shaped form. Two kinds of adhesive glands, the foot and style-glands as found in other planktonic rotifers, are recognizable at the coxal part of the toes, each measuring 12x7 µm in size. At the dorso-basal part of the toe, there is found one big caudal elevation, 10-20x4-8 µm in size. It is still quite uncertain whether or not these peculiar organs correspond functionally to the papillar organs found in some other parts of this animal. The egg produced adheres without exception to the caudal extremity of the mother without thread. (ref. ID; 3535)
Comments
A form notommatoformis occurs mainly in late spring, while form vermiformis occurs usually in mid-summer. And form tintinnoformis appears chiefly from late summer to autumn. (ref. ID; 3535)
Measurements
The amictic females of the present species are 72-152 µm in total length, 23-46 µm in height and 28-40 µm in width, and their eggs, 36-50x26-32 µm in size, are smooth without spines, folds or punctuations on the surface. The young just hatched are 64-72x23-24 µm in size and usually vermiform. The [male-female] females are 115-122x33-36 µm in size, their eggs are smooth and 30-31x22 µm in size; while the [D-female] females are 118x33 µm in size, their eggs produced during the period from September to November are similar to those of Trichocerca birostris (Sudzuki, 1957) in features, measuring 47-52x27-33 µm in size and often spiny. The present author could not decide if such two types of dormant eggs are simply referable to the difference of the species. The three sexual types of the females are hardly distinguishable by their size and outer appearance. The amictic females occur from the beginning of July to the beginning of December, the greatest maxima, lying mainly in September. The mictic females which appear about a week later than the amictic are found occasionally in small numbers during succeeding four months. (ref. ID; 3535)