The World of Protozoa, Rotifera, Nematoda and Oligochaeta
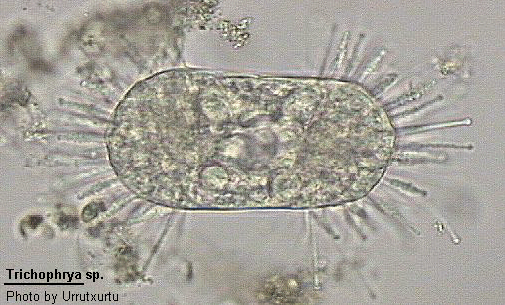
Trichophrya
Trichophrya Claparede & Lachmann, 1859 (ref. ID; 2013)
From Dr. Inaki
Class Kinetofragminophora: Subclass Suctoria: Order Suctorida: Suborder Endogenina (ref. ID; 2013)
[ref. ID; 2013]
Body irregularly spherical, directly attached to substratum. Capitate tentacles long and regularly arranged in fascicles. Neither stalk nor lorica present. Reproduction by simple or multiple endogenous budding producing an egg-shaped swarmer. Often reported growing upon the peritrich Epistylis. Macronucleus ovoid, elongate or branched but not coiled.
Most easily confused with Platophrya with which it is sometimes considered to be synonymous.
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013)
[ref. ID; 3672]
Species of the genera Trichophrya and Heliophrya are strikingly similar in structure, both being stalkless and dorsoventrally flattened with peripheral fascicles of tentacles. The genera are distinguished by their mode of budding, Heliophrya reproducing by evaginative budding and Trichophrya by endogenous budding. (ref. ID; 3672)
[ref. ID; 4282]
The genus Trichophrya was established by Claparede & Lachmann in 1859 and comprised several free-living and symbiotic species. Initially, Trichophrya was classified as a genus in the aloricate suborder of the family Acinetidae (Kent 1880-1881) but later placed in the family Dendrosomidae (Penard 1920) and ultimately in Trichophryidae (Corliss 1979; Gonnert 1935). According to the latest revised classification (Levine et al. 1980), Trichophrya is a genus within the suborder Endogenina. There is a need for some classification of a confusing literature on species of Trichophrya and Heliophrya, which show striking morphological similarities in that both genera are stalkless, dorsoventrally flattened, and have tentacles in fascicles. The genus Heliophrya, established by Saedeller & Tellier (1930) and also named Craspedophrya (Rieder, 1936), belongs to the newly established family Heliophryidae in the suborder Evagogenina (Corliss 1979). The mode of embryogenesis therefore provides a distinction between Trichophrya and Heliophrya. (ref. ID; 4282)
[ref. ID; 3672]
Trichophrya collini has a polygonal, or slightly irregular, dorsoventrally flattened cell body (45-75 um diam.), attached to the substratum by an adhesive disc. A 170-nm-thick rim is present around the dorsal periphery. The capitate tentacles are up to 500 um long and arranged in fascicles located on slight protrusions at each angle around the periphery. There are up to 15 tentacles within each fascicle, arranged in 1-3 rows, and held at an angle of up to 90 degrees to the cell surface. The apical knob of the tentacle has protuberances marking the position of internal haptocysts. The tentacles are highly sensitive to mechanical simulation, reacting by contraction. A prominent, central (12-15 um diam.), oval or kidney-shaped macronucleus with distinct chromatin bodies is adjacent to a 1.5-2-um-diam. micronucleus. A contractile vacuole (diam. 1.5-2 um) is located on either side of each fascicle. (ref. ID; 3672)
Cell body: The 0.1-1-um-thick fibrous adhesive disc extends up to 5 um beyond the cell body. The dorsal and ventral cortex differ in thickness and complexity. The dorsal cortex has a 0.2-6-um-thick external coat, which is absent on the ventral side. The underlying plasma membrane and two inner membranes cover the entire cell except the tentacle, apical knob, and the pore lumina, where only the plasma membrane persists. A granular layer (8-12 nm thick) occurs beneath the dorsal membranes, but it is reduced or absent ventrally. The innermost layer, the epiplasm, is filamentous, 50-70 nm thick dorsally, and 25-50 nm ventrally. Both the dorsal and ventral cortex penetrated by pores. The body cytoplasm contains food vacuoles, membrane-containing bodies, crescentic bodies (particularly in recently fed cells), and 0.7-1.2-um Gram-negative bacterial endobionts. Membrane-bounded elongate (90-100x200-760 nm) dense bodies and haptocysts 510 nm long are also widespread but are most common around the tentacle base. The contractile vacuole complexes consist of a vacuole lumen, surrounding spongiome, and a ventral discharge canal. (ref. ID; 3672)
Tentacle: The tentacles display the typical suctorian pattern but also have some unusual features. The apical knob, which is stellate in section, has haptocysts with the distal end embedded in a 12-nm-thick electron-dense material adjacent to the plasma membrane. In some micrographs, their proximal ends are connected to a capsule within the apical knob by rods 25 nm in diameter. The haptocysts have the usual shaft, collar, and bulb but also have three groups of radial projections in their collar region. The axoneme is attached to an electron-dense rim just below the apical knob and runs deep into the body cytoplasm. The axoneme consists of an outer ring made up of six groups of four single microtubules, each group being linked to one of the six curved lamellae (each made up of five linked microtubules) that form the inner ring. Some lamellar microtubules bear inwardly pointing arms. Within a 700-850-nm region immediately below the knob, the axoneme is significantly wider (420-550 nm); only a few outer ring microtubules persist, and the lamellae are arranged in a helix. Below this region, the outer ring microtubules, increased here to 28 or 29 in number, are not separated into groups, but are joined on their inner side by a 5-nm-thick connective sheath. Near the proximal end of the axoneme, only a few of the outer ring microtubules persist. (ref. ID; 3672)
Reproduction: A single equatorially ciliated oval swarmer 30-35 um long is produced by endogenous budding. The dorsal cortex in vaginates to form a brood pouch with active cilia. The pouch enlarges to include part of the body cytoplasm within the developing swarmer. Towards the end of swarmer formation the macronucleus divides unequally, the smaller portion being incorporated into the swarmer. The swarmer remains, sometimes rotating, within the brood pouch for about 30 min. Embryogenesis takes 3.5-4 hr from the time of invagination. The liberated swarmer swims for 20-30 min before settling. Metamorphosis at first involves cessation of ciliary movement, ciliary loss, and cell expansion. Knobbed tentacles begin to appear at distinct points along the periphery within 50 min of settling, and a polygonal body shape is established. Initially, one tentacle develops at each angle, but soon (60 min) a second arises to establish the first row within the fascicle. Within less than 3 hr from the start of metamorphosis, an adult cell with 5-6 tentacles within each fascicle has been formed. (ref. ID; 3672)
Comments
[ref. ID; 3672]
Although T. collini, like T. rotunda, shows differences in complexity between ventral and dorsal cortex, these are not so pronounced as in H. erhardi, where the two inner membranes are absent ventrally. The significance of the central location of the contractile vacuole discharge canal exit pore in both T. collini and T. rotunda is unknown. (ref. ID; 3672)
Reproduction: Trichophrya collini resembles T. rotunda in producing a single equatorially ciliated swarmer by endogenous budding. Metamorphosis is relatively rapid in T. collini (3-3.5 hr) as compared with the 24 hr of T. rotunda. In T. collini, the first tentacles are equally spaced around the periphery of the disc-shaped young cell, and it is possible that the subsequent polygonal outline is caused by the enlarged tentacle bases developing within the fascicle. Unlike D. collini, the tentacles in T. collini are knobbed from the time of their first appearance, as in Acineta tuberosa. Trichophrya collini is thus equipped for prey capture in as little as 1 hr after settling. (ref. ID; 3672)
Systematics: Evidently, T. collini, previously referred to as Heliophrya collini, has been assigned to the wrong genus. Although the two genera are similar in appearance, there are, as indicated above, clear ultrastructural differences. More particularly, T. collini does not show evaginative budding. The difference in the mode of reproduction between the two genera is reflected in the difference in the size, shape, and ciliation of the swarmer. That of T. collini is oval with equatorial ciliature and resembles the swarmer of T. rotunda rather than the flattened swarmers with ventral ciliature of H. erhardi and other evaginative budders. Although T. collini and T. rotunda show morphological and ultrastructural similarities, they are distinct species. In T. collini the body is polygonal in shape; the shape is emphasized by a peripheral rim, which is absent in the more rounded T. rotunda. The tentacles of T. collini are always arranged in distinct fascicles at the body angles, but in T. rotunda fascicles may be absent. The tentacles within a fascicle are arranged in rows as P. rotunda, C. rotunda, and H. collini, unlike the irregular arrangement in T. rotunda. The tentacle of T. collini lacks the proximal connective sheath of T. rotunda. The swarmer of T. collini is distinctly larger than the swarmer of T. rotunda but similar in size to that reported for P. rotunda and H. rotunda. Metamorphosis is more rapid in T. collini than in T. rotunda. (ref. ID; 3672)
Body elongate, irregular or rounded; cytoplasm often filled with yellow to orange spherules; a single micronucleus; a single contractile vacuole; attached to the gill of small mouth black bass, Micropterus dolomieu. (ref. ID; 1618)
Measurements
Body up to 30-40 um long by 10-12 um; fully extended tentacles 10-12 um long. (ref. ID; 1618)
[ref. ID; 4282]
Trichophrya rotunda has a round (37-73 um diam.) oval-shaped, or irregular, dorsoventrally flattened body attached to the substratum by an adhesive disc 2.5 um in diameter. The tentacles are randomly arranged within each of from four to thirteen fascicles (sometimes indistinguishable). Each fascicle has one to six 37-190x1 um capitate tentacles, which project from the periphery at an angle of 15-45 degrees and thus exceed the cell body diameter by a factor of three to four. The tentacle knob has closely packed protuberances marking the position of the internal haptocysts. The cell body contains a prominent polymorphic but unbranched 2.5x12.5 um macronucleus with a closely associated micronucleus and granular cytoplasm. There are 7-21 peripheral contractile vacuoles, each 1-4 um diam. (ref. ID; 4282)
Cell body: The adhesive disc is about 0.16 um thick and composed of fibers 10 nm in diameter forming a loosely woven network, which often includes debris and bacteria. The cortex consists of an external surface coat, a triple-layered membrane system, a granular layer, and an epiplasmic layer with associated microtubules. There is as distinct difference between the dorsal and ventral cortex, the latter being thinner and less complex. The dorsal cortex possesses a 0.2-0.5 um thick external coat, which ventrally is reduced or absent. The triple-layered membrane system consists of a plasma membrane covering the entire cell body and two inner membranes which are absent from the tentacle knob and lumens of the cortical pores. The electron dense granule zone beneath the membranes is 20-27 nm thick dorsally but is reduced ventrally. The innermost layer, the fibrous epiplasm, is prominent dorsally (95-155 nm thick) but again is reduced ventrally. The cortex is penetrate by pores. The peripheral contractile vacuoles are surrounded by spongiome and have permanent discharge canal as observed inner suctoria. The canal is surrounded by helically wound microtubules and radial microtubules are associated at its proximal end. Of particular note is the ventral position of the discharge pore. The body cytoplasm contains the usual protozoan organelles and also a substantial number of crescentic bodies, food vacuoles, strong bodies, membrane-containing bodies, and two types of oval or elongate (0.4x0.9 um) bacterial endosymbionts. The macronucleus contains numerous granules (17 nm diam.) in the nucleoplasm, round or elongated (0.1-1.3x0.3-2.2 um) chromatin bodies with a fibrous substructure, and bundles of microtubules. The micronucleus contains a granular nucleoplasm, little condensed chromatin material, and is situated near the macronucleus. (ref. ID; 4282)
Tentacle: The tentacles are highly sensitive to mechanical stimulation, reacting by contraction. They also display a high degree of flexibility, bending movements being especially evident during feeding. In the tentacle cytoplasm, two specifically suctorian organelles, elongate dense bodies (EDB) and haptocysts, are abundant. The 150-650 nm, membrane-bounded EDB tend to be accumulated at the base of the tentacle, within the axoneme lumen, and in the center of the apical knob. The 400 nm long haptocysts occur particularly around the tentacle base, within the periaxonemal space, and embedded in the plasma membrane of the apical knob. The common three main regions of the haptocyst -shaft, collar, and bulb- are evident. Of particular interest are unusual radial projections originating from two crescent-shaped pads of the collar region. The projections of the distal pad point downwards while those of the proximal point upwards. The axoneme exhibits a typical suctorian fine structure but with some unusually features. It runs from its attachment immediately below the apical knob to deep into the body cytoplasm where it may follow a curving or twisting course. The prevailing microtubule pattern throughout the axoneme length is that of two concentric regions, an outer ring (0.5-0.65 um diam.) of 35 single microtubules, some of which may be interlinked, and an inner ring of seven curved lamellae, each of six closely linked microtubules bearing a short arm directed towards the lumen. The outer ring, however, is separated into seven groups of five microtubules with each group being directly linked with a component of its corresponding lamella. A number of modifications of this prevailing pattern are present. Immediately below the apical knob, the lamellae splay out and are set in a helix, only a few of the outer ring microtubules persisting at this point. About 2 um below the attachment point, the outer ring microtubules, now increased to 37, are no longer separated but are joined on their inner side by an 8 nm thick connective sheath. A similar modification is evident proximally just above point of axoneme termination in the body cytoplasm. Here the outer ring microtubules are joined on their inner side by a basal connective sheath. Finally, at the point of axoneme termination in the body cytoplasm the lamellae splay out and only a few single, possibly outer ring, microtubules persist. (ref. ID; 4282)
Feeding: Trichophrya rotunda readily feeds on and even shows a preference for vorticellid telotrochs. More common suctorian prey organisms, such a Colpidium, are readily captured and ingested but the suctorian does not survive. Capture of larger prey ciliates, such as Paramecium, has not been observed. Up to three cells may be ingested simultaneously. The prey may exceed the suctorian in size by a factor of two or three, but the vorticellids give little resistance to capture. Ingestion is completed within 1.5 hr with a final discarding of the residual prey. In all the feeding animals observed, there was attachment of the knob only at the trochal band. Additional feeding tentacles even underwent significant bending in becoming attached to these regions. (ref. ID; 4282)
Reproduction: Budding is endogenous, producing a single swarmer within a brood pouch. The first sign of reproduction in the light microscope is an invagination of the dorsal cortex. Formation of a birth pore is followed by that of a brood pouch in which active cilia of the incipient swarmer are evident. Gradually the pouch enlarges, carving out part of the adult cytoplasm to be enclosed within the swarmer. Towards the end of swarmer formation, the macronucleus divides following a change in the chromatin material from elongated bodies to strand-like threads. Following macronuclear division, the bridge connecting the swarmer and the parent's cytoplasm gradually disappears, completing swarmer formation. The parent macronucleus remains next to the brood pouch during the remaining part of reproduction. The separated swarmer remains within the brood pooch for about 20 min during which time it undergoes rotatory movements. It then moves to the birth pore and is squeezed out. From TEM it is evident that kinetosome replication takes place prior to formation of the brood pouch. Prokinetosomes are formed perpendicular to barren kinetosomes within a circular field set in finely granular material. The swarmer (10-17 um in diameter) is slightly flattened and oval-shaped. Its cytoplasm contains no tentacle axoneme primordia nor any of their developmental stages. The cell is rich in ribosomes, rough ER, and EDB. Crescentic bodies, fully formed haptocysts, and electron-dense bodies are evident throughout the cytoplasm. The macronucleus contains condensed chromatin material, nucleoli, and bundles of three to six microtubules. The swarmer has five equatorial ciliary rows. The cilia have the common 9+2 structure with doublet arms and radial spokes. Proximally each kinetosomes has the typical 'cartwheel' arrangement while distally the A tubules are joined by an inner 4 nm electron-dense band and ciliary necklaces are evident. The kinetosome-associated fibers include basal microtubules of unknown origin running underneath the kinetosome at right angles to its axis and a transverse fiber as defined by de Puytrac (1970), arising from triplets 3 and 4 and running to the left of the kinetosome and towards the surface. A kinetodesmal fiber originates from triplets 6 and 7 and runs to the right, but the ultimate fate of this fiber is unknown. Finally, three postciliary microtubules run from triplets 8 and 9 perpendicular to the basal microtubules. A parasomal sac occurs on either side of the kinetid. (ref. ID; 4282)
Comments
Trichophrya rotunda was cited as a new species by Hentschel (1915) but has since been described under the names of Platophrya rotunda (Gonnert, 1935), Craspedophrya rotunda (Rieder, 1936), and most recently as Heliophrya rotunda (Matthes, 1954). Morphological resemblances to H. collini (Saedeleer & Tellier, 1930) are evident also. Hentschel (1915) descried T. rotunda as a round (70 um diam.), stalkless cell with the tentacles arranged in irregular peripheral groups (often indistinguishable), a kidney or heart-shaped macronucleus, and 7-21 peripheral contractile vacuoles. The available descriptions thus make the systematic validity of T. rotunda and H. rotunda questionable and it is evident that the existing classification of species in the families Trichophryidae and Heliophryidae is in need of some revision. (ref. ID; 4282)
Systematics
It is evident that T. rotunda (Hentschel), most recently renamed Heliophrya rotunda by Matthes (1954), does not belong to that genes. A 'Heliophrya'-like ultrastructure is absent and, most significantly, it does not show evaginative budding. Although their morphologies are similar (Dragesco et al., 1955), ultrastructural differences between T. rotunda and H. erhardi include the cortex, haptocysts, bacterial endosymbionts, and the tentacle axoneme. The tentacle of T. rotunda shows distinct differences in both thickness and substructure to that of H. erhardi (Hascal & Rudzinska 1970; Lanners 1978; Spoon et al. 1976). The T. rotunda cortex does, however, resemble that of T. epistylides. The radiating projections of the haptocysts are also absent in H. erhardi but present in T. epistylides as well as T. rotunda. The tentacle axonemes of the two genera show distinct differences. Trichophrya rotunda (and T. epistylides) have an axoneme with the microtubules of the outer cylinder arranged in fives and with apical and basal connective sheaths. Heliophrya erhardi (Hauser & Van Eyes, 1976) has axonemes with no subdivisions in the microtubules of the outer cylinder and with no basal connective sheath but it does have a basal fibrous collar. The most important difference between the two, the mode of reproduction, is emphasized by the differences in the size, shape, and ciliation of the swarmer. Clearly the organism described here is not of the genus Heliophrya, but of Trichophrya; but it is dissimilar to the H. collini described by Saedeleer & Tellier (1930), C. rotunda by Rieder (1936), and H. rotunda reclassified by Matthes (1954). This dissimilarity is based on the lack of organization into tentacle rows within a fascicle and on the observation that distinct fascicles are not always present in this organism as they are in H. erhardi, C. rotunda and H. rotunda. The suctorian studied here best matches the description for T. rotunda given by Hentschel (1915) and there is therefore a need to re-erect Trichophrya rotunda (Hentschel). (ref. ID; 4282)