The World of Protozoa, Rotifera, Nematoda and Oligochaeta
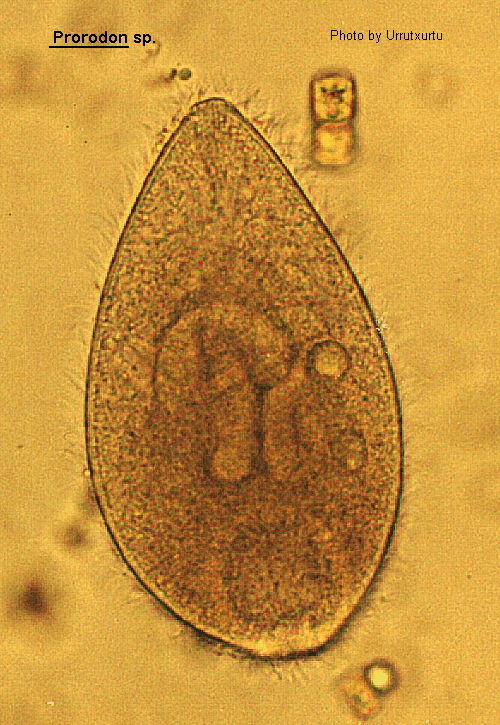
Prorodon
Prorodon Ehrenberg, 1833 (ref. ID; 2013, 4611, 4881, 7273)
From Dr. Inaki
Class Kinetofragminophora: Subclass Gymnostomata: Order Prostomatida: Suborder Prorodontina: Family Prorodontidae (ref. ID; 2013)
Family Prorodontidae Kent (ref. ID; 4813)
Synonym Rhagadostoma Kahl (ref. ID; 1618)
Subgenus Rhagadostoma Kahl, 1926 (ref. ID; 3540)
[ref. ID; 2013]
Body shape ovoid to cylindrical, in some species the body narrows posteriorly. Oral aperture circular, apical, leading to a cytopharynx which is supported by large, distinct double trichites. Somatic ciliation uniform, in longitudinal meridians whose kinetosomes may be paired. The meridians with 1 or 2 exception reach the apical oral aperture. There are 3 parallel double rows of kinetosomes above the short meridians and these comprise the basal bodies of the dorsal 'brush'. A few species have a tuft of caudal cilia. Macronucleus rounded to elongate. One or more contractile vacuoles in posterior region. Some species feed upon protozoa and algae, others are histophagous.
See Pseudoprorodon.
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013)
[ref. ID; 3596]
Body ellipsoid or stout to elongate ovate, oval in cross sections; anterior end usually rounded, sometimes oblique; posterior one round, somewhat convex conical to a blunt end; mouth terminal or subterminal at the anterior end, conveyed to a rod-shaped pharynx; macronucleus spherical, ovoid, or ribbon-like; contractile vacuole posterior, often single; cilia uniform except in the oral region. (ref. ID; 3596)
[ref. ID; 4757]
Foissner (1983) proposes the following diagnoses for the genus Prorodon (translated into English): "Prorodontidae with a meridionally orientated long dorsal brush to which somatic kineties of one side of the body abut under an acute angle." To allow the distinction between the "dexiotrop" and the "aristerotrop" type the diagnosis should be modified as follows: "Prorodontidae with a meridionally orientated long dorsal brush to which somatic kineties from the left half of the cell body (seen from outside of the cell) abut upon the brush under an acute angle." The genus Paraprorodon defined by Foissner (1983) would represent the "syntrop" type shown in Fig.13. Foissner's diagnosis: "Prorodontidae whose dorsal brush is formed by wavy kineties which are followed by a file-shaped organelle" cannot be accepted because P. aklitolophon also shows a file-shaped structure. According to our SEM observations a similar structure is also seen in those representatives which belong to the "dexiotrop" type. A wavy appearance of the brush is also seen in silver stained preparations of the "dexiotrop" group, therefore we recommend to neglect this trait. We suggest the following modified diagnosis for the genus Paraprorodon Foissner, 1983: "Prorodontidae with a dorsal brush to which somatic kineties abut on from both sides, the right and the left side." Consequently, separate genera should be created for the "enklitoloph" and the "enklitoloph - aristerotrop" group. We have observed certain ultrastructural features which point to a close homogeneousness of the Prorodon groups shown in Fig.13. Moreover these 4 groups differ distinctly from the other genera of the family Prorodontidae Kent, 1881 as defined by Corliss (1979). In particular some of the genera carrying the word "prorodon" in their names, like Spiroprorodon Fenchel and Lee, 1972, Pseudoprorodon Blochmann, 1866 and Helicoprorodon Faure-Fremiet, 1950 differ clearly from the 4 Prorodon subgroups. We, therefore, would like to see the taxon Prorodontidae restricted to the 4 subgroups illustrated in Fig.13. For the Prorodontidae the following new diagnoses is suggested: "Holotrichous ciliates with an apical or subapical cytostome and meridionally arranged somatic kineties. There are 3 meridionally arranged kinetofragments which cover 1/5 to 1/2 of the body length in immediate neighbourhood to the cytostome." For P. parafricanus Dragesco, 1986 which is the only Prorodon to show a brush made of four kineties, an electron microscopical investigation would be desirable to check whether this species really belongs to the Prorodontidae. If this is really the case, the diagnosis for the mainly in question will have to be changed again. The remaining genera -Bursellopsis Corliss, 1960; Helicoprorodon Faure-Fremiet, 1950; Pseudoprorodon Blochmann, 1866; Spathidiopsis Fabre-Domergue, 1889 and Spiroprorodon Fenchel and Lee, 1972- of the Prorodontidae sensu Corliss (1979) should -as far as it has not been done already [see Foissner (1978) and Small and Lynn (1986)]- be separated from the Prorodontidae sensu stricto. If these view is accepted nothing would stand against establishing 4 genera for the groups shown in Fig.13. Before taking this step we rather recommended to wait for the further development in the systematics of the "prostome" ciliates, which seem to be good for more surprises. (ref. ID; 4757)
[ref. ID; 4881]
The genus Prorodon Ehrenberg, 1833 currently holds more than 60 nominal species. These include aerobic and anaerobic ciliates (Esteban et al. 1993) from freshwater and marine environments. There is a general consensus (Corliss 1979; Small & Lynn 1985; Hiller & Bardele 1988) that ciliates in this genus are medium to large holotrichous ciliates, ballon-shaped, with an apical or subapical cytostome, meridional somatic kineties, and a "brosse" (brush hereafter) with three files of dikinetids to which the somatic kineties may abut from either side of the cell. Caudal cilia are present in some species. The typical life cycle is a succession of morphologically diverse vegetative forms, including a division and a resting cysts (Hiller & Bardele 1988). This polymorphism may be the root cause of the inflated number of species in the genus. Silver impregnation techniques are usually successful with Prorodon, and the species can be distinguished using three main conservative features: (1) morphology and arrangement of the brush (see Hiller & Bardele 1988); (2) number and arrangement of the somatic kineties; and (3) morphology of the oral structures. (ref. ID; 4881)
Prorodon aklitolophon Hiller & Bardele, 1988 (ref. ID; 4757 original paper, 7273) reported author and year? (ref. ID; 191)
Diagnosis
P. aklitolophon is a small, relatively slender representative of the genus Prorodon. The theront measures 75 um x 25 um and has 35 meridional somatic kineties. Its brush consists of 3 likewise meridional kinetofragments, 20 um in length which are not touched neither by the right nor by the left somatic kineties. This means that both, the somatic and the brush kineties show a parallel orientation. One single shortened somatic kinety ends at the posterior end of the leftmost brush kinety. The length of the apical cytopharyngeal basket measures 20 um. Near the posterior end of the cell there is a conspicuous pulsatile vacuole. Moreover, there are 4-16 stiff caudal cilia almost twice the length of the ordinary somatic cilia. The macronucleus is kinety-shaped, 20 um x 8 um in size and normally located in the middle of the cell. A single micronucleus (2.4 um x 1.5 um) lies close to the macronucleus. (ref. ID; 4757)
Life cycle
[ref. ID; 4757]
P. aklitolophon is a histophagous ciliate which passes through distinct developmental stages during its life cycle. The various stages of the life cycle are so different in size and shape that they may easily be regarded as belonging to different species unless their continuous transformation is observed. The terms used in this paper for the different life cycle stages (theront-trophont-protomont-tomont-tomit) were introduced by Mugard (1948) for Ophryoglena but are now widely used in the description of other histophagous ciliates, too. Depending on the availability of food two cycles may be distinguished in P. aklitolophon: the division cycle when there is plenty of food and the starvation cycle when food is scarce or not available at all. (ref. ID; 4757)
The division cycle: Asexual reproduction in P. aklitolophon takes place in division cysts only. Theronts freshly released from such cysts measure 75 um x 25 um and swim around vividly searching for food. Rounded at both ends the cell has the shape of a slightly bowed cylinder. When swimming the anterior end of the cell describes a cone-shaped path while the entire cell moves forward in a clockwise motion parallel to its longitudinal axis, covering about ten times its body length per second. Kept at 18 degrees C young, hungry theronts swim around for about 30 hr before they settle down to form a resting cysts. If during this period food is added they immediately start feeding and within 2-3 hr these stated (now called trophonts) change into protomonts. Theronts seem to be attracted to the food by some chemosensory mechanism. Well-fed protomonts measure 100 um by 65 um and are elliptical in shape. The interior of the protomont is filled with numerous spherical food vacuoles. Thus the protomont appears less translucent than the theront. Based on the average size of these two stages a 4.4-fold increase in volume is calculated for the transformation of the theront into the protomont. Parallel to the increase in size the swimming motion of the protomonts slows down. Finally they settle down and it is within 3-6 hr after the beginning of feeding that they have formed division cysts (tomonts) usually in group of 5 to a couple of hundreds at the wall of the culture dish. At 18 degrees C the further development to the mature theront ready to escape from the cyst again takes 3.5-4 days. At 20-22 degrees C the development takes 2-3 days and at 10 degrees C 12-14 days, respectively. In P. aklitolophon the number of the tomites formed per division cyst is not invariably fixed but depends on the size of the tomont. Thus in tomonts of 45-75 um in diameter 4 tomites are formed while in tomonts of 30-60 um only 2 tomits are formed. Tomonts with more than 4 tomits were never observed. (ref. ID; 4757)
The starvation cycle: If the theronts find no food within 1-3 days after the release from a division cyst they encyst forming a resting cyst of their own. The "first generation resting cyst" measures about 40 um in diameter and, apart from its smaller size, it is easily distinguished from a division cyst by its higher transparency. Contrary to the tomont the cell in a resting cyst stays ciliated. The length of the period a starving cell spends in the resting cyst is highly variable. While some cells reappear as free-swimming theronts after a few hours others may stay in their cysts for several days. If a starving theront finds no food within a certain period of time it encysts again and the individual cell may repeat the starvation cycle several times without cell division. During continuous lack of food the period spent in the resting cyst becomes longer, while at the same time the period spent as free-swimming theront becomes shorter. The size of the starving theronts becomes smaller and smaller, their cytoplasm becomes more translucent and vacuolated and the swimming speed is reduced. After a period of continuous starvation for 4-8 weeks (!) the resting cyst measures 25-35 um in diameter and the starving theronts are 40 um by 15 um. Being of such small size they are now unable to feed and finally die. The formation of dormant cysts to overcome longer periods of starvation without intermitting encystment was never observed no matter at which temperature the starving cells were kept. (ref. ID; 4757)
The theront: The theront represents the fully differentiated, free-swimming stage in the life cycle of P. aklitolophon. Due to the absence of food vacuoles this stage is best suited for the description of the species-specific characters and it can also most easily be prepared for light and electron microscopic observations. The true shape of the free swimming cell can only be observed with the dissecting microscope, since any microscopical technique which demands a coverslip will result in an artificial outline of the cell due to compression. The oral funnel does not lie exactly at the apex of the cell but is slightly shifted to the ventral part of the left side of the cell. It is laterally compressed, thus distally showing a slit-like opening with slightly curved rim. Usually the anterior half of the cell looks more transparent than the posterior half. In slightly squeezed cells the cytopharyngeal basket is clearly seen in the anterior third of the cell. In Protargol-stained specimens the cytopharyngeal basket seems to consist of some 30 rods. In living as well as in glutaraldehyde-fixed theronts 3 very conspicuous parallel kinetofragments, about 20 um in length, are seen to lie between the somatic kineties. This structure, known as "dorsal brush", almost reaches the oral slit. At the posterior end of the dorsal brush there is one single shortened somatic kinety (post-brush kinety). It is the only somatic kinety which does not reach the oral opening. The somatic ciliature of the cell, which is most clearly seen in stained preparations, consists of about 35 kineties with some 45 kineties. Close to the oral opening the cilia appear to be more closely spaced and bent towards the oral funnel. These cilia measure 5 um in length and are thus shorter than the other somatic cilia which measure 8 um in length. In living specimens the large pulsatile vacuole, which is a characteristic feature for all members of the genus Prorodon, is clearly visible. When squeezed cells are observed with phase optics, water-collecting channels running parallel to each kinety are seen to be a regular component of the pulsatile vacuole. These collecting channels taper off towards the anterior end of the cell thus coming into close contact with the microtubule system of the somatic kineties. Near their posterior end they show a distinct ampullalike widening close to the point where they are connected to the proper pulsatile vacuole. The finer details of this water expulsion system are seen best in living cells which are still able to perform the rhythmical systole - diastole cycle. Such a highly evolved drainage system showing a close association with the somatic ciliature has, as far as we know, not yet been described for any of the supposedly primitive gymnostomes. This drainage system seems to be characteristic for the genus Prorodon in general, as we have found it also in P. teres, P. raabei and another not yet determined Prorodon species. The fine structural details of this Prorodon - specific drainage system will be subject of a separate contribution (Hiller & Bardele in preparation). P. aklitolophon has a single kidney-shaped macronucleus (20 um x 8 um) with a distinct nucleolus and a single micronucleus (2.4 um x 1.5 um), both nuclei are lying in the middle of the cell. At the posterior pole of the cell there are 4-16 stiff caudal cilia 14-16 um in length. (ref. ID; 4757)
Comments
This newly described species has been cultured in the laboratory for now more than 2 years under different culture conditions. The morphology of all stages of the life cycle as well as the variation of all relevant biometrical data have been compared with other hitherto described species. As a result none of the hitherto described Prorodon species seemed to be identical with our species, so we had to describe it first before starting our comparative study of the genus with the final intention to settle the question of the phylogenetic evolution of the so called gymnostome - prostome holotrichs. These are 4 other Prorodon species which show a similar orientation of the somatic kineties on both sides of the brush as P. aklitholophon. These are P. discolar (Jordan 1974), P. gracilis (Groliere 1976), P. ovalis (Dragesco 1970) and P. pyriformis (Groliere 1976). There is no necessity to discuss all Prorodon species described so far, instead we concentrate on those 4 species just mentioned which show a similar array of the somatic kineties in close neighbourhood of the dorsal brush as observed in P. aklitolophon. The stage of life cycle which is best suited for the diagnosis of the species-specific characteristic is the free-swimming stage which occurs in all species whether as theronts or permanently free swimming-stages. To differentiated these 5 (or actually only 4) species the infraciliature, the number of somatic kineties, the approximate number of kinetids per kinety, the number of post-brush kineties and the number and relative length of the caudal cilia can be used (needless to say that these characteristics in combination with the life cycle are also most valuable for classifying the other species of the genus Prorodon). We have to point out that the drawing of the theront of P. discolar has been modified compared with the original publication by Jordan (1974). This author has drawn a theront in which the somatic kineties on both sides of the triple-rowed brush run parallel to it, this is not in agreement with the published micrographs, where left of the brush 7-10 kineties are seen to abut on it in an acute angle (see also Foissner 1983). We thus conclude that P. discolar has a similar arrangement of the somatic kineties on the left side of the brush as e.g. P. teres (enklitoloph-dexiotrop brush). A similarity of P. discolar with P. aklitolophon can therefore be excluded. The 3 other species (P. gracilis, P. ovalis and P. pyriformis) do have an "aklitoloph" brush. The number of the somatic and the post-brush kineties, the absence or existence of the caudal cilia are sufficient to differentiate P. ovalis and P. pyriformis from P. aklitolophon.
During our studies of P. aklitolophon we realized that the structural organization of the brush in connection with the adjacent somatic kineties is a conspicuous trait which can easily be seen by means of almost any kind of silver impregnation. The term used are taken from the Greek and have the following meaning: "..." (aklitoloph) = kineties not inclined upon the brush, "..." (enklitoloph) = kineties inclined upon the brush. The term "..." (dexiotrop), "..." (aristerotrop) and "..." (syntrop) are necessary to differentiate the 3 possible types of an "aklitoloph" brush; their meaning is "reaching towards the brush from the right, the left or both sides", respectively. Within these 4 groups the qualities partly mentioned already, may be used to characterized the particular species. Most important characteristics are the number of the post-brush kineties and the number of the kineties abutting the brush. According to our experience the variability of these characteristics is fairly small and can easily be handled by biometrical analysis. Together with the appearance of the caudal cilia, the comparison of the silverline patterns and, if applicable, the difference in life cycle, a well-defined characterization of the various Prorodon species should be possible. (ref. ID; 4757)
Etymology
The name for the new Prorodon species refers to a character which is easily seen with the light microscope and which distinguishes this species from many other species of the same genus. This character is the course of the somatic kineties in relation to the brush kineties. The fact that the somatic kineties are not inclined to the brush was the reason why the Greek name ... (aklitolophon) as chosen (meaning: "not leaning to the brush"). (ref. ID; 4757)
Ovoidal; 45-55 ciliary rows; macronucleus ellipsoid; micronucleus hemispherical; contractile vacuole terminal; fresh water. Kahl (1930) states that it occurs also in brackish water containing 2.5 per cent salt; sapropelic form in salt water is said to possess often long caudal cilia. (ref. ID; 1618)
Body large, worm-like, 2.6-3.8 of the breadth in length, rounded in the anterior end, broadest in the suboral region, then gradually narrowing towards a blunt posterior end; mouth terminal, pharynx funnel-shaped; macronucleus spherical, central; contractile vacuole indistinct; fine yellowish granules scattered in the protoplasm. (ref. ID; 3596)
Comments
The species differs from P. teres Ehrenberg, P. marinus Claparede & Lachmann, and P. moebiusi Kahl in larger size and in the presence of yellowish granules and from P. morgani Kahl in the form of the macronucleus which is spherical in this species, but elongated in the latter. This species seems to be the longest among species of Prorodon, and resembles P. moebiusi in general contour. If the size of the body and the colour of granules are variable in this form, this species is probably identified with P. moebiusi. (ref. ID; 3596)
Body ellipsoid; mouth opening terminally with an elongated conical pharynx; macronucleus oval with a micronucleus, having a clear area in the center; contractile vacuole distal; cilia of the oral margin growing forwards and those of the posterior region somewhat longer (ref. ID; 3596)
[ref. ID; 7273]
Light microscopy: The theront stage of P. teres measures approximately 160x70 um. About 90 meridionally arranged somatic kineties cover the cell body. The anterior part of each kinety consists of 5 to 12 dikinetids followed by approximately 140 monokinetids comprising the remaining posterior portion of the row. The parasomal sac adjacent to the dikinetids is also stained in the Fernandez-Galiano preparations. The somatic cilia are about 10 um long with the cilia of the anterior dikinetids being slightly shorter. At the posterior end of the cell, there are 8-16 long, stiff caudal cilia (ca. 20 um long).
The outer rim of the cytostomial funnel, which is situated slightly to the side of the apical pole of the cell, can be recognized in Fernandez-Galiano preparations as a darkly stained circle consisting of 40-50 paired nematodesmata. These rods are about 45 um in length and extend slightly obliquely into the cell. The brush of P. teres is of the dexiotrop type (Hiller and Bardele 1988) and covers about one-fourth of the cell length. The brush is composed of three rows of dikinetids. About 14 somatic kineties abut its right side in an acute angle. The first three somatic kineties (K1 to K3) on the right side of the brush are close together for the first half of their length. This feature is unique to P. teres (Foissner 1983). Further characteristics of P. teres are the numerous 12-15-um-long needle-shaped and slightly curved toxicysts and a sliverline system with very small mesh size. At the posterior pole of the cell near the cytoproct are several large pores of the contractile vacuole. Furthermore, each cell shows many smaller contractile vacuole pores randomly distributed in the posterior third of the cell. In a few preparations of P. teres a subpellicular infraciliary lattice can be seen.
The nuclear apparatus of P. teres, usually situated in the center of the cell, consists of a spherical to ovoid macronucleus (32x24 um) and a spherical to lenticular micronucleus (8.5x4.7 um) in close contact to the macronucleus.
Prorodon teres is histophagous, with a life cycle very similar to that of P. aklitolophon (Hiller and Bardele 1988). Cell division occurs in cysts and always results in two tomites. Therefore the difference in volume between theronts and protomonts in P. teres is less conspicuous than it is in P. aklitolophon, which undergoes two division cycles to give four tomites. (Hiller and Bardele 1988). In the various stages of the life cycle the ratio of length:width varies between 1.7 (protomonts) and 2.4 (theronts). The slimmest and longest forms correspond to the recently released theronts whereas starved theronts become shorter and, because their width does not decrease dramatically, they look rounded. (ref. ID; 7273)
Electron microscopy:
Pellicular alveoli and ectoplasm. Immediately underneath the cell membrane, P. aklitolophon and P. teres show an elaborated system of pellicular alveoli. Within the alveoli numerous dense granules can be observed, which, in this pronounced form, are characteristic for the genus Prorodon. In tangential sections the pellicular alveoli appear as a more or less polygonal network with a mesh size of 1-1.5 um. Normally one cilium is surrounded by three pellicular alveoli. The height of the pellicular alveoli varies, measuring 2.5 um in the anterior parts of P. teres (1.5 um in P. aklitolophon), and normally decreases toward the posterior pole of the cell. In transverse sections, the inner alveolar membrane appears to be irregularly curved, following the surface of a dense layer of epiplasm with a thickness of about 15 nm. The epiplasm itself is folded, forming circular ridges of varying diameter, which give the inner surface of the pellicular alveoli its characteristic shape. That is the reason why tangential sections running at the level of thee circular ridges show at first glance a confusing pattern of pellicular alveoli, epiplasm, cytoplasm, extrusomes (mucocysts and toxicysts) and cilia. Each cilium is surrounded by one circular cytoplasmic ridge. The ridges of consecutive cilia are fused on the right side of each kinety, forming a continuous cytoplasmic ridge. In this ridge, extending as a more or less straight line from posterior to anterior, two to five longitudinal microtubules can be found. These microtubules are sandwiched in the distal tip of the ridge between the epiplasm and the inner alveolar membrane.
All extrusomes (mucocysts and toxicysts) lie between the kineties. They are not grouped and seem to be more or less evenly distributed all over the cell with an accumulation of toxicysts close to the cytostomial funnel. In contrast to this, stacks of flattened vesicles can be regulary observed in the close vicinity of the parasomal sacs. Just underneath the epiplasm is a filamentous network. The microfibrils forming this network are aggregated into thick bundles with a diameter of 40-80 nm, reaching up to 120 nm in P. aklitolophon and 200-300 nm (maximally 600 nm) in P. teres. Kinetosomes and extrusomes, which extend deeper into the cytoplasm, are surrounded by these microfibrils. (ref. ID; 7273)
Comments
P. teres is usually found in fresh or brackish waters, while the allied species, P. marinus, is known as a sea form, but the specimens are characteristic in the structure of the oral border and the macronucleus, hence the identification has been made. (ref. ID; 3596)
Identified by large contractile vacuole that occupied at least one third of the body which measured 75x25 um. Large oval macronucleus present. In some individuals posterior region distinctly spatulate. (ref. ID; 2316)
P. viridis is a medium-sized ciliate, typically 70-90 um in length. Its brush is formed by three parallel rows of dikinetids which extend to the equator of the cell. Somatic kineties usually number 35-40, and all but one reach the oral aperture. The oral aperture is lined with inconspicuous cytopharyngeal rods. There is a single post-brush kinety. Caudal cilia are present. The macronucleus is single, spherical or ovoid, with one adjacent micronucleus. Symbiotic zoochlorellae are always present in the cytoplasm. This species has a characteristic life cycle, with a permanently planktonic, vegetative stage, and with cell division by binary fission in the water column. (ref. ID; 4881)
Descriptions
The organisms are ellipsoidal, sometimes almost spherical, typically 70-90 um long and 50-80 um wide. Cell length can vary from 45 to 140 um depending on the physiological stage and on the size and number of ingested prey. Consequently, the cell shape too is variable; it can be spherical, or truncated at the anterior end and pointed at the posterior, or nearly cylindrical. The diet of the ciliate is extraordinarily diverse and related to what is available and abundant in the water column. Food items include rotifers and their egg, gastrotrichs, diatoms, euglenids, cryptomondas, dinoflagellates, other heterotrophic flagellates, and ciliates. The ciliate has been observed to eat the corpses of Euglena. Food vacuoles are frequently absent in small cells of P. viridis, although zoochlorellae are always present. Three conservative features identify P. viridis: (1) the presence of symbiotic zoochlorellae in the cytoplasm; (2) the long brush formed by three parallel kineties, with one post-brush kinety of variable length and, (3) the inconspicuous cytopharygeal rods (nematodesmata) lying beneath the oral dikinetids. The oral area, brush, and symbiotic Chlorellae were briefly characterized by Finlay et al. (1988). Silver-impregnation (Chatton-Lwoff, and Fernandez-Galiano methods) of both wild and cultured organisms, reveals a fairly constant pattern of infraciliature. The number of somatic kineties is 35-39, infrequently up to 43, plus the short post-brush kinety. All somatic kineties on both sides of the brush reach the oral aperture although in some organisms kinety 2 is slightly shorter and finishes instead at the dorsal brush. There is a variable number of kinetosomes per kinety, from 25 to more than 50, the number being correlated with cell size. The post-brush kinety varies in length too, from 2 to 20 kinetosomes (most frequently 15), but this variation is independent of cell size. Recently divided organisms can show morphological variations, such as more than one somatic kinety ending at the brush and not reaching the oral area, variable length or presence of more than one post-brush kinety, four rows in the brush, and complete absence of the brush. The brush is typically formed by three parallel rows of kinetosomes that originate at the oral aperture and always extend to the cell equator. In silver-impregnated organisms, each of the rows seems to be formed by paired kinetosomes, one of which is ciliated. The oral infraciliature is formed by dikinetids with microtubular ribbons. Other morphological features observed in impregnated cells are the single contractile vacuole pore, which opens at the rear of the organism, and the darkly-stained extrusomes visibly distributed over the entire surface of the cell. There is a single spherical macronucleus with one adjacent micronucleus. Post-conjugation organisms with two or three micronuclei are common. The caudal cilia are easily observed in living organisms, and they usually number four to ten. (ref. ID; 4881)
Life cycle: The striking characteristic of P. viridis is its planktonic life style and the permanently free-swimming stage of the vegetative form. Unlike most other Prorodon species, P. viridis never divides in a cyst, and the different stages of the life cycles -theronts, trophonts, and protomonts- are not easily discriminated. Organisms begin morphogenesis independently of their size, and the symbiotic zoochlorellae are present in all stages of the life cycle. To our knowledge, only on other species of Prorodon, P. multinucleatus (Dragesco 1960), has been reported with cell division outside of a cyst. The infraciliature of this species, however, was not provided and its allocation to the genus Prorodon is not yet definetively established. (ref. ID; 4881)
Remarks
P. viridis bears some resemblance to two other prorodontids, i.e. P. aklitolophon (Hiller & Bardele 1988), and Bursellopsis (Hiller 1991). However, P. aklitolophon never has endosymbiotic Chlorellae and its life cycle involves division and resting cysts (Hiller & Bardele 1988). Ciliates in the genus Bursellopsis (e.g. Bursellopsis spaniopogon Hiller, 1991), do divide outside of a cyst, and the oral infraciliature resembles that of P. viridis, but the somatic infraciliatures are distinctly different, e.g. the long kinetodesmal fibres in Bursellopsis spp. are never found in P. viridis. Historically, only two "green" Prorodon species have been reported more than once: these are P. ovum and P. viridis. Until recently (Foissner et al. 1994) no descriptions based on silver staining were available for either species. P. ovum was first described by Ehrenberg (1831) as Holophrya ovum, with symbiotic zoochlorellae. Kahl (1935) did not mention zoochlorellae in P. ovum, but he had previously (1927) described two new species of green ciliates, P. viridis and P. minutus. In 1994 Foissner et al. described P. ovum with silver impregnation techniques, and synonymized P. ovum and P. viridis as Holophrya ovum. Simultaneously, these authors transferred the P. ovum described by Augustin & Foissner (1992) to Holophrya discolor. Many other workers have consistently found two non-overlapping zoochlorellae-bearing species of Prorodon, e.g. Agamaliev (1986), in freshwater and brackish water lagoons in the Caspian Sea, Alekperov & Kryuchkov (1982) in a fish hatchery in Azerbaijan; Czapik (1975) in Crakow; Dragesco & Dragesco-Kerneis (1986) in Benin and Cameroon (Africa); Pedros-Alio et al. (1995) in Lake Ciso (Spain). In conclusion, it appears that two different zoochlorellae-bearing Prorodon "morphospecies" exist (see Finlay et al. 1996) - both are cosmopolitan, but they differ in size and infraciliature. The larger species (120-160 um) should be referred to as P. ovum (Ehrenberg, 1831) and smaller (50-120 um) as P. viridis Kahl, 1927. The latter absorbs P. minutus Kahl (1927), and P. deflandrei Dragesco (1960). (ref. ID; 4881)