Dileptus
Dileptus Dujardin, 1841 (ref. ID; 2013, 4861)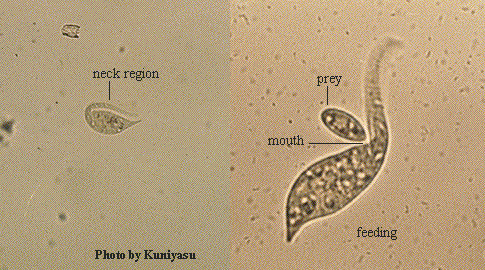
Body shape highly elongate, size ranges 100-1,600 um long, rounded in cross-section, with long highly mobile, contractile prehensile anterior neck region. Ventral surface of neck lined with trichocysts. Some species have a pointed tail region, others are rounded posteriorly. At the base of neck lies the oral aperture which is supported by a cytopharyngeal basket of trichites (not always easily visible). The neck is often held extended, bent towards the dorsal surface. Trichocysts commonly present in cytoplasm and particularly down the unciliated ridge of the neck. Ciliation complete, in form of longitudinal kineties. Contractile vacuoles usually numerous in row along dorsal surface often with large terminal vacuole. Contractile vacuoles rarely on both dorsal and ventral surfaces. Macronucleus highly variable from species to species but usually in 2 to many parts which may be distributed throughout the cytoplasm or may be moniliform, rarely single but when so is elongate. Originally described by Dujardin (1841).
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013) [ref. ID; 4861]
Recent investigations showed that the genus Dileptus is polyphyletic: D. amphileptoides apparently lacks a usual pharyngeal apparatus (Foissner 1984) and D. monilatus lacks oblique preoral kineties (Foissner et al. 1995). In fact, years ago these species were chosen by Jankowski (1967) as types for two new subgenera of Dileptus (with many macronuclear nodules), viz. Dimacrocaryon (with two macronuclear nodules) and Monilicaryon (with moniliforme macronuclear), using their macronuclear configurations as sole diagnostic charcter; an unfortunate action because the same macronuclear pattern very likely evolved independently in several evolutionary lines of Dileptus s. l. Foissner (1984) thus amended Jankowski's diagnosis of Dimacrocaryon and suggested uniting binucleate species with a normal oral apparatus in a new genus, Rimaleptus. However, he did not appreciate this genus for the reason mentioned above. Further studies are necessary to elucidate whether or not the macronuclear of configuration can be used to split the genus according to its evolutionary wheer or not the macronuclear configuration can be used to split the genus according to this evolutionary history. For the present, it seems wise to keep all species which possess a usual infraciliature in the time-honoured genus Dilpetus. The somatic and oral infaciliature of most Dileptus species is very similar, except for the new species mentioned above and possibly some others. Thus, the classical species characters (size, shape, length of proboscis, nuclear apparatus, number and location of contractile vacuoles, biotope) used by Kahl (1931) and Dragesco (1963) are still valid. However, I would like to add another highly useful character, viz. the extrusomes which show a bewildering, yet hitherto unrecognized diversity in size, shape and arrangement (Song Weibo et al. 1988, present results and data from about 10 new terrestrial species not yet published). Usually these organelles do not stain with protargol and their shape is more or less altered by various fixatives. Thus, size, shape and arrangement of the extrusomes must be studied in live cells. (ref. ID; 4861)
- Dileptus aculeatus Dragesco, 1960 (ref. ID; 3690 original paper)
- Dileptus alpinus Kahl, 1931 (ref. ID; 1622, 2128, 3593)
- Dileptus americanus Kahl, 1931 (ref. ID; 4861) reported year? (ref. ID; 1618, 1622)
- Dileptus amphileptoides Kahl, 1931 (ref. ID; 3593) reported author and year? (ref. ID; 1622)
- Dileptus anatinus Golinska, 1971 (ref. ID; 4861) reported author and year? (ref. ID; 4612)
- Dileptus anser Dujardin, 1841
See; Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683) - Dileptus anser (O.F. Muller, 1786) (ref. ID; 1219, 1335, 1622, 2245, 2571, 3115, 3593, 4683) reported year? (ref. ID; 1618, 3698) reported author and year? (ref. ID; 3292, 4612, 4686, 7596)
Syn; Amphileptus anser Ehrenberg, 1838 (ref. ID; 3115); Amphileptus cygnus Claparede & Lachmann, 1859 (ref. ID; 4683); Amphileptus margaritifer Ehrenberg, 1838 (ref. ID; 3115); Dileptus gigas f. grojecensis Wrzesniowsky, 1870 (ref. ID; 4683); Dileptus gigas f. varsaviensis Wrzes., 1870 (ref. ID; 1622); Dileptus irregularis Maskell, 1888 (ref. ID; 1622); Vibrio anser O.F. Muller, 1786 (ref. ID; 4683) - Dileptus beersi (ref. ID; 4861)
- Dileptus binucleatus (ref. ID; 1622)
- Dileptus bivacuolatus Da Cunha, 1915 (ref. ID; 1622, 4488)
- Dileptus conspicuus Kahl, 1931 (ref. ID; 3593) reported author and year? (ref. ID; 1622)
- Dileptus costaricanus Foissner, 1995 (ref. ID; 4861 original paper)
- Dileptus cygnus (Claparede & Lachmann, 1859) (ref. ID; 1622, 2571) reported author and year? (ref. ID; 3292)
- Dileptus dubius Vuxanovici, 1959 (ref. ID; 4861)
- Dileptus elephantinus Svec, 1897
See; Paradileptus elephantinus (Svec, 1897) Kahl, 1931 (ref. ID; 4612, 4613) - Dileptus estuarinus Dragesco, 1960 (ref. ID; 3690 original paper)
- Dileptus falciformis Kahl, 1931 (ref. ID; 3593) reported author and year? (ref. ID; 1622)
- Dileptus fasciola Fromental, 1874
See; Litonotus fasciola (ref. ID; 3115) - Dileptus gigas (Claparede & Lachmann, 1859) (ref. ID; 1622, 2245)
Syn; Phragelliorhynchus nasutus Herrick, 1884 (ref. ID; 1622) - Dileptus gigas f. grojecensis Wrzesniowsky, 1870
See; Dileptus anser (ref. ID; 4683) - Dileptus gigas f. varsaviensis Wrzes., 1870
See; Dileptus anser (ref. ID; 1622); Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683) - Dileptus gracilis (ref. ID; 1622)
- Dileptus irregularis Maskell, 1888 (ref. ID; 4683)
See; Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683) - Dileptus lacazei (Gourret & Roeser, 1886) (ref. ID; 1622)
- Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683)
Syn; Amphileptus anser Ehrenberg, 1838 (ref. ID; 4683); Amphileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683); Dileptus anser Dujardin, 1841 (ref. ID; 4683); Dileptus gigas f. varsaviensis Wrzesniowsky, 1870 (ref. ID; 4683); Dileptus irregularis Maskell, 1888 (ref. ID; 4683) - Dileptus margaritifer (Ehrenberg, 1833) Dujardin, 1841 (ref. ID; 4612) reported author and year? (ref. ID; 1629)
Syn; Amphileptus margaritifer Ehrenberg, 1833 (ref. ID; 4612) or 1838 (ref. ID; 3115) - Dileptus marinus Kahl, 1933 (ref. ID; 1620) reported year? (ref. ID; 3690)
- Dileptus massutii Kahl, 1933 (ref. ID; 1620)
- Dileptus monilatus (Stokes, 1886) (ref. ID; 1622, 1896) reported year? (ref. ID; 1618) or (Stokes, 1886) Kahl, 1931 (ref. ID; 4612)
See; Monilicaryon monilatus (ref. ID; 4612) - Dileptus mucronatus Penard, 1922 (ref. ID; 1622, 4522, 4861)
- Dileptus orientalis Song Weibo et al., 1988 (ref. ID; 4861)
- Dileptus robustus Vuxanovici, 1959 (ref. ID; 4861)
- Dileptus saaleri Schwarz, 1962
See; Pelagodileptus trachelioides (ref. ID; 4613) - Dileptus similis Foissner, 1995 (ref. ID; 4861 original paper)
- Dileptus tenuis Penard, 1922 (ref. ID; 1622, 3593)
- Dileptus terrenus Foissner, 1981 (ref. ID; 4861)
- Dileptus thononensis Dragesco, 1960 (ref. ID; 3690 original paper)
- Dileptus trachelioides Zacharias, 1894 (ref. ID; 4613) reported year? (ref. ID; 2226)
See; Pelagodileptus trachelioides (ref. ID; 4613) - Dileptus viridis (Ehrenberg, 1833) (ref. ID; 4730)
Syn; Amphileptus viridis Ehrenberg, 1833 (ref. ID; 4730) - Dileptus visscheri Dragesco, 1963 (ref. ID; 7689) reported year? (ref. ID; 3698)
Dileptus americanus Kahl, 1931 (ref. ID; 4861) reported year? (ref. ID; 1618, 1622)
Descriptions
Proboscis bent dorsally sickle-like; macronucleus made up of 2 sausage-shaped or often horseshoe-shaped parts; 2 contractile vacuoles on dorsal side; in mosses. (ref. ID; 1618)Measurements
200 um long. (ref. ID; 1618)Dileptus anser (O.F. Muller, 1786) (ref. ID; 1219, 1335, 1622, 2245, 2571, 3115, 3593, 4683) reported year? (ref. ID; 1618, 3698) reported author and year? (ref. ID; 3292, 4612, 4686, 7596)
Synonym
Amphileptus anser Ehrenberg, 1838 (ref. ID; 3115); Amphileptus cygnus Claparede & Lachmann, 1859 (ref. ID; 4683); Amphileptus margaritifer Ehrenberg, 1838 (ref. ID; 3115); Dileptus gigas f. grojecensis Wrzesniowsky, 1870 (ref. ID; 4683); Dileptus gigas f. varsaviensis Wrzes., 1870 (ref. ID; 1622); Dileptus irregularis Maskell, 1888 (ref. ID; 1622); Vibrio anser O.F. Muller, 1786 (ref. ID; 4683)Descriptions
Slender with neck-like elongated proboscis at the anterior end which is slightly flattened and highly contractile; cytopharynx exhibiting long trichocysts; uniform ciliation all over the body, 2 rows of strong cilia and 3 rows of trichocysts along the ventral side of the proboscis; macronucleus divided into 200-500 small bodies; about 20 micronuclei; numerous contractile vacuoles along the dorsal side. (ref. ID; 1219)Proboscis slightly flattened; macronucleus divided into 100 or more discoid bodies; sixteen or more vesicular micronuclei; contractile vacuoles in row on the aboral surface, with two to three in proboscis; in fresh water. (ref. ID; 1618)
Measurements
Length 200-600 um, average about 350 um. (ref. ID; 1219)250-400 um, sometimes up to 600 um long. (ref. ID; 1618)