The World of Protozoa, Rotifera, Nematoda and Oligochaeta
Synchaeta
Synchaeta Ehrenberg, 1832 (ref. ID; 7815)
Class Monogonontate: Order Ploimida: Family Synchaetidae (ref. ID; 6806)
ref. ID; 822
This genus was erected to comprise three pelagic, essentially freshwater, illoricate rotifers, S. pectinata Ehrenberg, S. oblonga Ehrenberg, and S. tremula (O.F. Muller). Since 1832, 41 additional Synchaeta species have been described as new and several of these have been formally divided into subspecies or varieties. Approximately 36 of the nominal 44 species are generally recognized as valid. About 40% of the approximately 36 recognized Synchaeta species have been encountered in brackish water or marine habitats. Eighteen species could be considered truly marine (normally occurring in salinities greater than 25 0/00 S); S. baltica, monopus, triophthalma, littoralis, cecilia, vorax, neapolitana, curvata, atlantica, johanseni, pachypoda, grimpei, tamara, glacialis, hyperborea, baccilifera, neapolitanoides, okai. (ref. ID; 822)
ref. ID; 1663
With four long, prominent sensory bristles on the corona. Body conical. Toes small. Mastax virgate. Planktonic species. (ref. ID; 1663)
ref. ID; 1923
Trophi virgate, with prominent V-shaped striated hypopharyngeal muscle, visible in ventral view of intact animal. Corona with auricles and setae. (ref. ID; 1923)
ref. ID; 2989
Identification of members of the Synchaeta genus can only be satisfactory when the animal is fully stretched out. (ref. ID; 2989)
ref. ID; 4591
Body more or less conical or bell-shaped, terminated into a foot with toes; but in a few cases the foot or toes are reduced. Corona convex, with four prominent sensory bristles; front furnished laterally with two ciliated auricles. Mastax virgate (rod-shaped). (ref. ID; 4591)
This species found marine. (ref. ID; 822)
Specimens examined had an elongated conical body different from the shape shown by Rousselet. The trophi show differences in the unci which were U-shaped with 5-6 lancet-shaped teeth in addition to the stronger tooth. A number of variations were also found, but since the material was preserved these may have been due to the action of the formalin which causes considerable changes in the form of most members of this genus. (ref. ID; 2281)
Synchaeta calva Ruttner-Kolisko, 1970 (ref. ID; 2603 original paper)
Descriptions
When the animal swims freely the general outline of the body is conical with the foot parallel-sided and clearly distinct from the rest of the body. Observed under a coverglass the body becomes more or less vase-shaped depending on the filling of the stomach and the presence or absence of an egg. The toes are small and pointed. The most remarkable feature of the new form is an apical field, completely naked and with a marked conical bulge in the ventral region formed by the protruding mastax. The four so called "styli" although characteristic for the rest of the genus could not be detected in any of the specimens examined; the ciliated apical tentacle and the eyespot are also missing. The dorsal antenna lies at the same level as the ciliated auricles; the lateral antennae situated at two thirds of the body length are minute and almost invisible. The big egg-shaped mastax lies just behind the mouth opening and causes the body wall to form the proboscis-like protuberance, which is very mastax is very often to be found outside the body. The mouth parts are of the type of the Synchaeta pectinata-stylata group having unci with a double pointed tooth only and the rami being simple plates. The oesophagus is thin, transparent and very long; the stomach lies together with the rest of the viscera in the hind end of the body giving to the animal the appearance of emptiness - as long as it does not carry an egg inside the body. The vitellarium contains 8 nuclei; the stomach glands are small. (ref. ID; 2603)
Comments
When I first saw a few individuals of the new form I thought they might belong to the species Synchaeta stylata in a similar way as the so called "Polyarthra proloba" belongs to other Polyarthra species different only in forming a sack-like prominence for the bigger mastax (Ruttner-Kolisko 1959). But the pattern of its independent occurrence and the striking absence of setae on the head - a feature which has hitherto been considered a genus characteristic of Synchaeta - makes it seem reasonable to name the new form. To emphasize its most important morphological feature I propose therefore the species name calva (bald-headed). There can be no doubt that Synchaeta calva belongs to the "Formenkreis" stylata-pectinata as opposed to the "Formenkreis" oblonga-tremula. Whether such a member of a Formenkreis is a valid species in the genetic sence is still an open question (Ruttner-Kolisko 1963, 1969). From the ecological point of view Synchaeta calva forms independent populations living in their own ecological niche and well defined by morphological structures. (ref. ID; 2603)
Measurements
Average of 10 adult living individuals) are: total length 250+/-5; max. width (between auricules) 130+/-5; foot 20+/-2; toes 10; mastax 65 µm. (ref. ID; 2603)
This species found marine. (ref. ID; 822)
The teeth on the unci were broader than usual and interlocked when the trophi were closed. In a few cases the usual cleft eyespot was separated into two distinct spots, but this may have been due to the action of the preservative. (ref. ID; 2281)
Marine and brackish water. (ref. ID; 3573)
Measurements
Total length of mature females in extended (swimming) posture 123-188 µm. (ref. ID; 822)
Overall length 100-123 µm. (ref. ID; 2281)
Marine. This is a distinctive species even in preserved collections. (ref. ID; 2281)
Comments
Rousselet (1909) separate bicornis from fennica on account of the additional frontal spots of the former, and the position and angle of the appendages. The last two characteristics would appear to be of little value, and specimens were examined, undoubtedly fennica, in which the position and angle of the appendages would fit either species. In two cases pigmented granules were found in the anterior median area, not sufficient, however, to be called eyespots. If, however, these were the rudiments of frontal eyespots there would appear to be no real difference between fennica and bicornis. (ref. ID; 2281)
This species found marine. It is a eurythermic, euryhaline species. (ref. ID; 822)
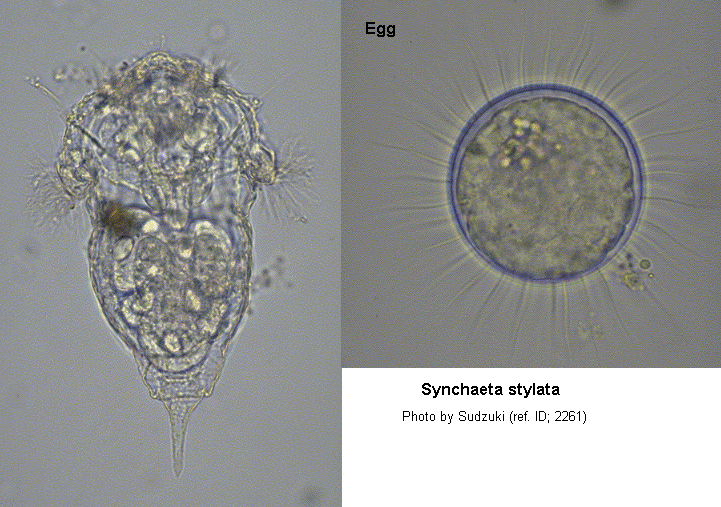
Female: Body of swimming individual is bell-shaped, fixed specimens are typically more bag-like. Subauricular appendices absent, although loose folds of integument form under auricles when the latter are extended. Single, large, variably shaped red eye of length (longitudinal direction) 13.5-17 µm and width 8.5-10 µm; typically well consolidated but sometimes with isolated pigment granules in periphery. Dorsal antenna present, consisting of about 4 filaments 12-15 µm long. Single lateral antenna on left side near foot base. Foot unsegmented, retractable, length 20-26 µm, bearing a single, sharply pointed, bilaterally symmetrical toe (length 10-12 µm), a bristle (spine?) (length 8.5-11 µm), originating on ventral side of foot and lying parallel to toe, and a spur (length 8.5-11 µm), projecting postero-dorsally from dorsal midline of foot. In life, the foot bristle is normally held against the toe and is extremely difficult to see. Of 100 formaldehyde-killed and stained (azo-black) individuals examined in succession on a microscope slide at x400, both the bristle and spur were discernible in 12: the bristle only was discernible in 17; the spur only in 27, and neither in 44. In these specimens the spur tended to be obscured because the feet were often partially retracted and nearly all the individuals presented only a dorsal or ventral view. Each uncus bears 6 teeth in addition to the sharply pointed structure located dorsally to the jaws. (ref. ID; 822)
Male: Not found. (ref. ID; 822)
Parthenogenetic egg: Ellipsoidal, major axis 65-70 µm, minor axis 60-65 µm. One or two eggs (very rarely three) are carried at a time. When a single egg is carried it is attached to the tip of the spur on the dorsal side of the foot. If a second egg is expelled it replaces the original egg on the spur tip and the latter is either dropped or loosely held in contract with the foot, body, and new egg. Sudden lethal heat shock causes eggs to detach; ovigerous individuals killed by cold formaldehyde solution usually retain their eggs. (ref. ID; 822)
Dormant egg: Not found. (ref. ID; 822)
Etymology
For colleague Dr. Larry Hutchings, oceanographer at the Sea Fisheries Research Institute, Cape Town. (ref. ID; 822)
Measurements
Total length of mature females in extended (swimming) posture 165-200 µm (mean 186); maximum width, including auricles 110-130 µm (mean 120), excluding auricles 85-100 µm (mean 92). Total length at hatching 85 µm; maximum width, including auricles 80 µm, excluding auricles 50 µm. (ref. ID; 822)
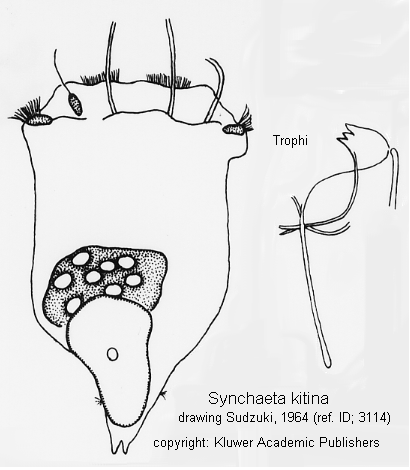
The lateral antennae were situated at the connection between body and foot. The lengths of the two toes were always equal. The animals examined had no finger-like projections under the 'Wimperohren' (Voigt 1957, p.396). The form in Blankvatn with these characteristics should be classified as S. tremula or S. kitina. These two species or sub-species (Sudzuki 1964) differ in body size as well as in number and form of teeth on the uncus. It was impossible to examine the uncus, but the size indicated that the form was S. kitina. Voigt (1957, p.400) gives the total length of S. kitina as 136 µm, and 177-222 µm for S. tremula. Sudzuki (1964) considers S. tremula as a sub-species of S. kitina giving in the names S. tremula kitina. (ref. ID; 2989)
Egg: Two different types of parthenogenetic eggs occur; (a) smooth-walled ones mainly in winter, and (b) spine-covered ones mainly in spring. The spined eggs are 55x60 µm to 65x75 µm and are slightly more rounded and larger than the smooth eggs. The ovarium has 8 nuclei and the animals have two bright red eye spots. (ref. ID; 1402)
This is typical coldwater species. The pentagonal corona is a good distinguishing mark, as it is often difficult to distinguish S. lakowitziana and S. oblonga if they co-occur; the mastax is of no great help, as even the unci are similar. The unci is two different type. The single teeth on the rami (reminiscent of those in P. dolichoptera) are conspicuous. The deep invaginations between the unci teeth differ from the figures of Lucks. The animals are usually deep yellow or brown, presumably because of their diatom diet. (ref. ID; 1850)
These forms were characterized by elongated body, pentagonal ("funfeckig") head and two semicircular eyes separate from one another. They also had two conspicuous constrictions, one behind the ciliated auricles and the other in front of the foot. (ref. ID; 3235)
Comments
Characteristics of S. lakowitziana, S. oblonga and S. truncata have been described. (1) S. lakowitziana is, as rule, decidedly bigger (ca. 250 µm) than the two other species, which are about the same size (ca. 150-200 µm). (2) Though the front end varies considerably in appearance with in each species, it is throughout more arched in S. lakowitziana and S. oblonga than in S. truncata, where it is relatively straight. On the other hand S. truncata does no always taper conically towards the hind end in the way shown by Rousselet (1902). In well-nourished individuals and during the formation eggs its body is often broadest in the middle (as in S. oblonga). (3) S. lakowitziana is more extended in length than the two other species. The two constrictions (behind the ciliated auricles and in front of the foot) are especially marked in this species. (4) The lateral antennae are situated farther back in S. truncata than in the other two species. However, they are usually very difficult to detect, especially in S. oblonga. (5) The entrails of S. truncata often have a yellowish tone, while they are almost colourless in the other two species. (6) Eggs (one or two) are often attached to the toes of S. oblonga and S. lakowitziana. Sometimes they are seen at a distance from the posterior end, attached to the animal by a thread of mucus. S. truncata was never seen to carry its eggs. (7) S. truncata has a tendency to attach itself to some support (e.g. to plants) and gyrate around the point of attachment. The other two species do not seen to have this habit. (8) In swimming, too, S. truncata exhibits more gyrating movements than S. oblonga, which mostly swims along smoothly in a quieter and more harmonious manner than the "nervous-looking" S. truncata. (S. lakowitziana was not studied intensively in this respect but ought on the whole to agree with S. oblonga). It is thus evident that the three species concerned differ distinctly, no only morphologically but also ethologically. Carlin (1943) says it is possible to distinguish S. lakowitziana from S. oblonga by studying the eyes. According to my experience, however, the characters he mentions are not altogether reliable. In S. lakowitziana the eyes are said to be rather far apart from each other. This may be the rule in preserved specimens, but in living animals the eyes are usually situated close together. According to Carlin the eyes of S. oblonga are unequal in size. This was actually the case in many (but not all) my specimens. In living animals two eyes of uniform size were often seen, and in addition a separate pigment body. When the individual contract, the latter joins one of the eyes which thus looks bigger than the other. Parise (1961) does not consider it possible to distinguish the species concerned by studying their mastax. The three species differ from one another in certain ecological respects. Thus S. lakowitziana is more sensitive to high temperatures than the other two, and during summer it only occurs in the hypolimnion of deeper lakes. The animal were moreover especially liable to die under the microscope. Occasionally S. oblonga and, especially, S. truncata were found in the epilimnion during the middle of summer, at temperatures between 15 and 20 degrees C. As a rule they were encountered in cold-water samples, however. S. truncata is not as decidedly planktic as the other two, constituting as edge species between the euplanktic and the benthic-periphytic environments. Its habit of attaching itself to plants and other supports, as well as its rather vigorous swimming movements are probably connected with this. Planktic rotifers usually carry their eggs, and this is the case in S. oblonga and S. lakowitziana but not in the semiplanktic S. truncata. (ref. ID; 3231)
Measurements
Total length (without the bristles at the anterior end) 270 µm. (ref. ID; 2553)
The body is stouter than in S. triophthalma, being more cylindrical in shape. There are two streams of purple granules connecting the frontal eyespots with the occipital. The lateral antennae are placed some three-quarters of the way down towards the foot, at the point where the trunk begins to narrow. (ref. ID; 1419)
Male: No males were observed at Plymouth. (ref. ID; 1419)
The general form of the head, its styles and sense hairs, the position of the lateral antennae, and the structure of the foot and toes, conformed largely with the description and reproduction by Rousselet (1902). A conspicuous character was the large cervical eye which is connected with two frontal eye spots by two streams of red granules. (ref. ID; 3235)
Marine and brackish water. (ref. ID; 3573)
Type locality
Originally described by Rousselet from specimens collected at Dundee by Hood. (ref. ID; 1419)
Measurements
The size of this species seems rather variable. Rousselet gives the maximum length as 238 µm. The specimens here depicted, and most of the other Leasowe specimens measured 192 µm or thereabouts, and the Plymouth Sound specimens agreed with this. (ref. ID; 1419)
This species found marine. It bears a spur on the dorsal side of the foot. This character suggests a close relationship between the S. neapolitana and S. hutchingsi. There is a fundamental difference between the two species in the number and placement of the lateral antennae. Additionally, S. neapolitana has a weakly or strongly bilobed eye, whereas the eye of S. hutchingsi is neither. (ref. ID; 822)
Marine species. (ref. ID; 3573)
Measurements
Total length of mature females in extended (swimming) posture 109-213 µm. (ref. ID; 822)
This species has only one, dark violet, eye spot. The animals are often pink coloured, particularly their anterior part. The eggs are similar to type II eggs of S. lackowitziana covered with fine spines. (ref. ID; 1402)
Measurements
Egg 65x80 µm to 70x85 µm. (ref. ID; 1402)
Body length 110-156 (contracted); widest part 98-110; head 58-60 long; foot+toes 38-40; fertilized dormant egg 50x50; egg spines 30-35 µm. (ref. ID; 3083)
Body length 190-290; body width 134-170; length of toes 35-50 µm. (ref. ID; 3275)
This species found marine. It bears single lateral antenna near the foot base on the left side, and lacks the spur. (ref. ID; 822)
This species is fairly easy to recognize, being extremely wide anteriorly, possessing, as a rule, three main pigment spots. A rather unique feature is the asymmetrical left lateral antenna, situated, not in the usual position (the lumber region), but on a prominent swelling on the left dorso-lateral surface of the foot. According to Rousselet, the right lateral antenna is absent, but the Hollowday has recently been informed by Mr A.L. Galliford that Remane has found what he believes to be the right lateral antenna in the normal position, but very inconspicuous, and without the brush of sensory setae. The foot terminates in a single pointed toe. (ref. ID; 1419)
Marine and brackish water. (ref. ID; 3573)
Type locality
S. triophthalma was originally found in 1893 by Dr. Lauterborn off the coat of Heligoland. (ref. ID; 1419)
Measurements
Total length of mature females in extended (swimming) posture 180-280 µm. (ref. ID; 822)
See S. lakowitziana
It gyrates in a characteristic way round its longitudinal axis being fastened by a thread from its foot to some suitable object. The toes were pointed and kept tightly pressed together, Also the truncate anterior end was clearly perceptible. (ref. ID; 3235)
Measurements
Total length 195 (on average) µm; the breadth between the auricles equalled 110 µm. (ref. ID; 3235)