The World of Protozoa, Rotifera, Nematoda and Oligochaeta
Keratella
Keratella Bory de St. Vincent, 1822 (ref. ID; 7815)
Class Rotatoria: Order Ploimida: Family Brachionidae (ref. ID; 7097)
Synonym Anuraea Ehrenberg, 1830 (ref. ID; 3514) or 1832 (ref. ID; 2978, 3688) reported year? (ref. ID; 1345, 4593)
ref. ID; 1663
Dorsal surface of lorica with a pattern polygonal facets. With one, two or no posterior spine and with six short to medium anterior spine. Highly variable plankton and limnetic species. Cyclomorphic. Very common species. (ref. ID; 1663)
ref. ID; 2891
The genus contains, to date, approximately 16 known species which are placed in recognized, defined morphological groups principally on the basis of the variable lorica form. There are three broad Formenkreis recognized: quadrata, valga-tropica and cochlearis, however there are as well several species groups which, although they show a certain morphological stability, are not, without further study, able to be ranked, e.g. reducta, lenzi, serrulata, americana, mixta-javana. The taxonomy of most polymorphic species at present, is based on morphological lorica markings, particularly the foundation pattern (facettation), the dorsal lorica, relative length of lorica spines and the surface structure. (ref. ID; 2891)
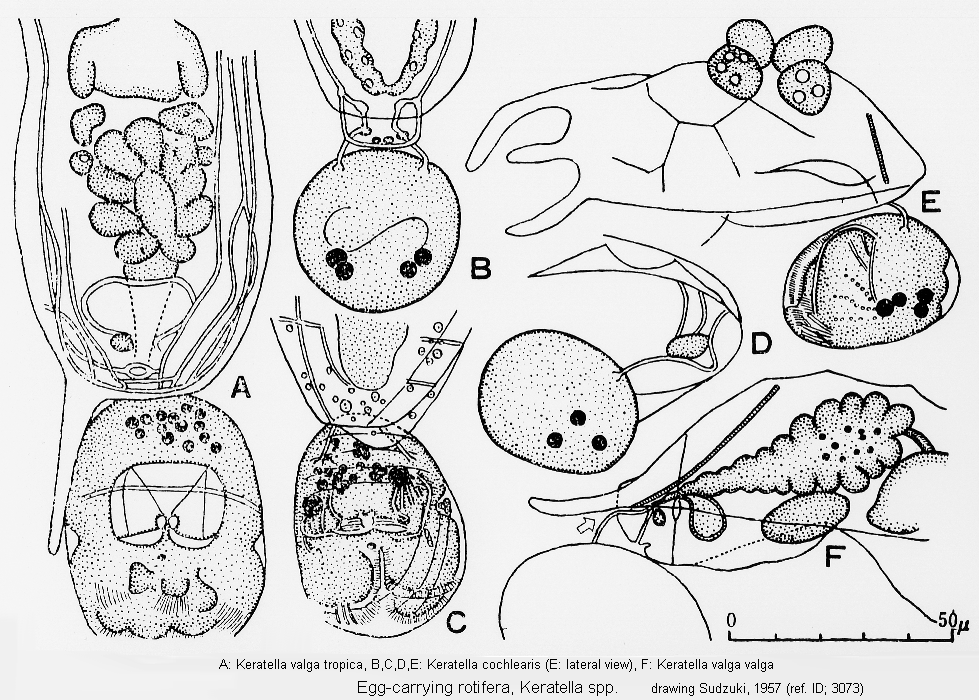
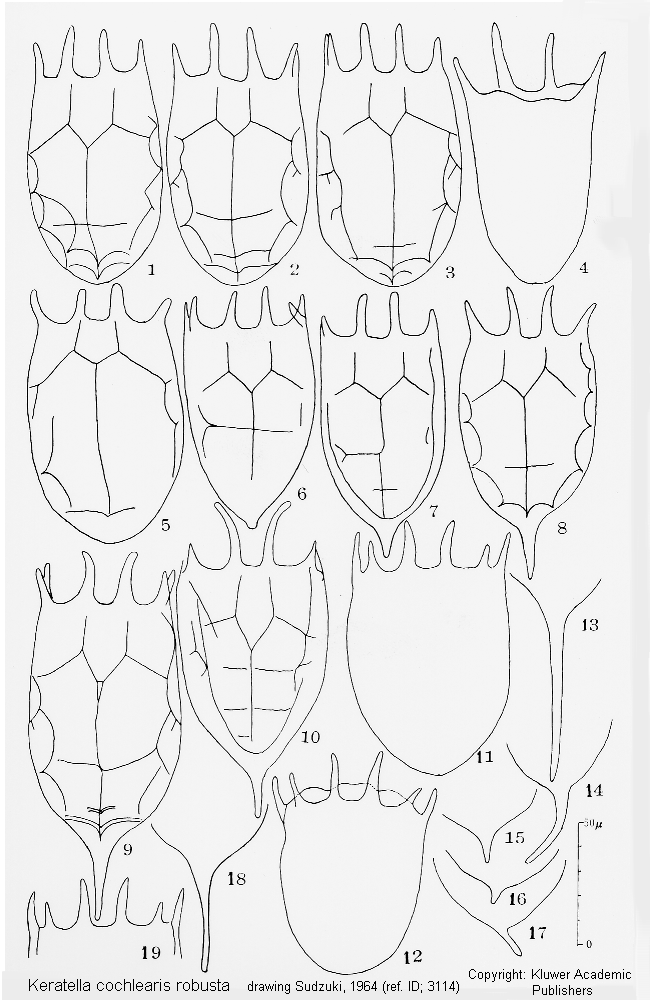
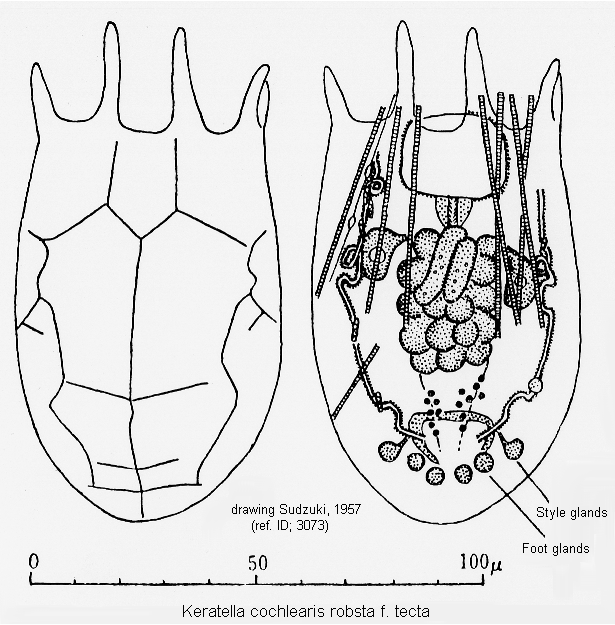
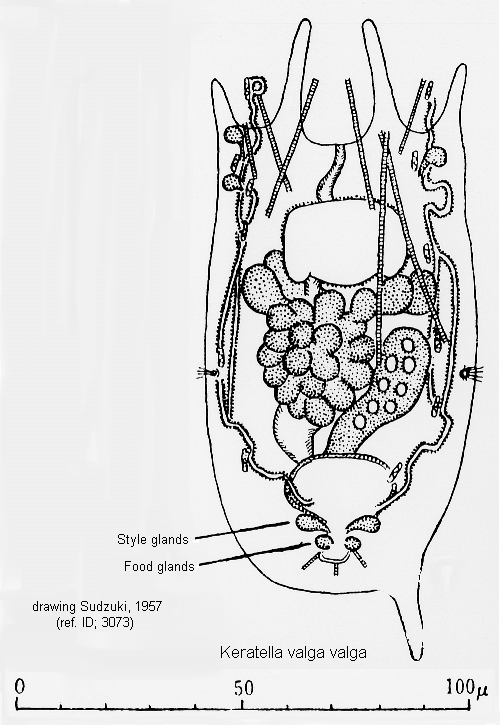
ref. ID; 3073
Mode of Egg-carrying (ref. ID; 3073)
Quote from ref. ID; 3073Quote from ref. ID; 3073
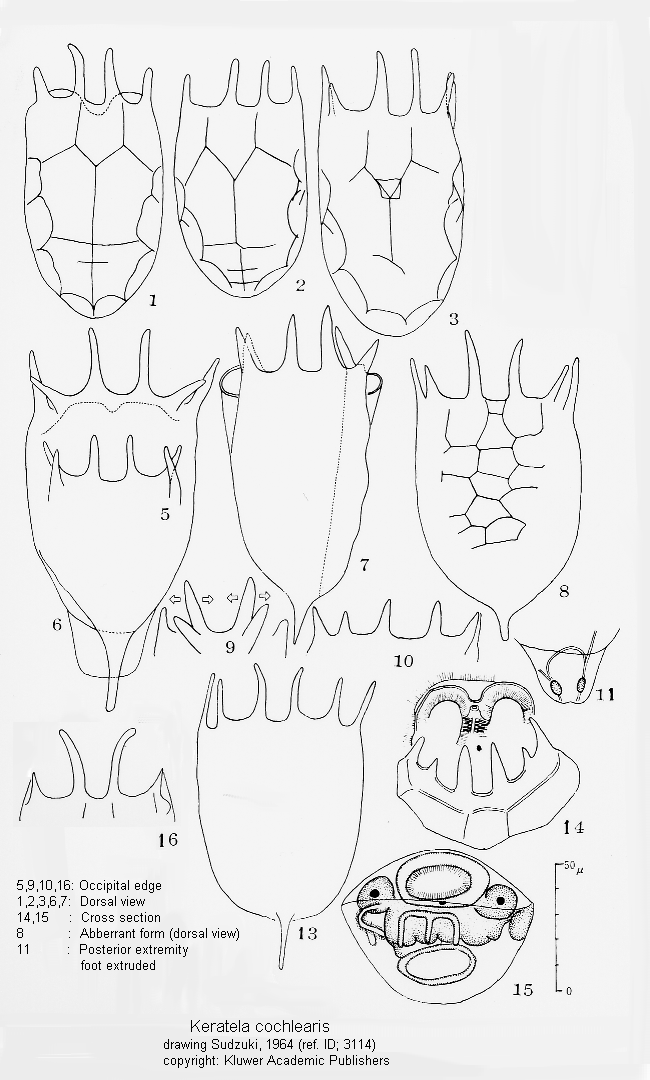
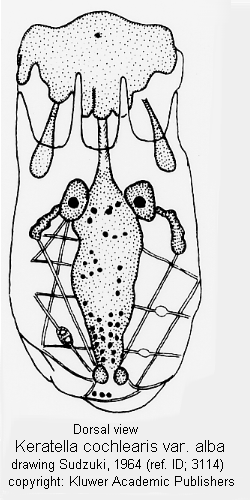
ref. ID; 3114
This genus is divided into several species based upon the following characters (Ahlstrom 1943; Gillard 1948): 1. The presence or absence of posterior spines, their number and position. 2. The pattern of dorsal sculpture on the lorica. 3. The proportions of the body, especially as regards size, shape, width and depth. 4. The number of anterior spines. 5. The ornamentation of the ventral plate. 6. The ornamentation of the dorsal plate (rather than the fundamental pattern of the dorsum). According to the Sudzuki's opinion the following characters are available for classification of the species. 1. Position of the posterior spines (terminal for cochlearis group, lateral for quadrata-valga group). 2. Features of the dorsal sculpture on the lorica (as the difference between americana and cochlearis, cochlearis and irregularis, serrulata and cochlearis, or testudo and quadrata). 3. General shape (or proportions) of the lorica (as the difference between K. valga and K. quadrata, cochlearis and faluta, or pyriformis and valga). 4. Features of the occipital spines (as the difference between taurophala and cochlearis). Such characters as presence or absence of the posterior spines, the ornamentation (except for sculpture) of the dorsal plate, however, have no taxonomic value. The number of anterior spines is so constant in this genus contrary to Brachionus, that it might serve for generic separation when it is different (cf. K. reducta). On the contrary, certain dorsal sculpture seems to be a good character for 'subspecific' distinction (as the difference between K. valga valga and K. valga tropica or between edmondsoni and quadrata or irregularis and earlinae). (ref. ID; 3114)
ref. ID; 4593
Brachionid rotifers with loricate body, separated into a dorsal and ventral plate. Dorsoventrally ore or less compressed. Anterior margin usually with six spines. Posterior spines often present, one or two in number, single posterior spine usually median. Dorsal plate with sculptural pattern, with or without areolate network, postulation, or fine spines. Many species in fresh water, only a few in brackish water. Keratella cruciformis cruciformis, cr. eichwaldi, quadraa platei, and cochlearis recurvispina are typical brackish-water forms. (ref. ID; 4593)
Keratella aculeata valga Rodewald, 1935
See; Keratella valga (ref. ID; 3688)
Keratella cochlearis and related species have very important problem of their morphological variability and possible transitions between different forms. Lauterborn (1900, 1904) has put forward his theory explaining different forms of the Keratella cochlearis group by step-by-step changes in morphology and splitting of a uniform winter Keratella into a number of successively separating strains of forms during summer, all of then ending up in the winter form again at the end of the season. Apart from minor objections and corrections the theory is still generally accepted in Europe, although it has never been proven experimentally. According to Lauterborn, the three main strains deriving from the winter form, Keratella cochlearis fa. typica are: a) the "macracantha-micracantha-tecta" strain with all cochlearis characters, but a successive reduction of body dimensions and an accelerated reduction of the posterior spine, b) the "hispida" strain, with an increasing pustulation of the lorica along with a successive disappearance of the plaque pattern, and c) the "irregularis" strain with the development of an additional "irregular" plaque, interrupting the median ridge of the lorica. Even in the "hispida" and the "irregularis" strains, the posterior spine is successively reduced. Besides the variable strains, a non-variable, "robusta" group may be found throughout the year, with basically the typical cochelaris characters.
Ahlstrom, on the basis of many thousands of careful measurements on material from all over the continent, from Mexico to Alaska, and comparison also with material from Europe and other places, calls into question the direct connection between K. cochlearis and the hispida and irregularis strains respectively. While he accepts, the definition of the robusta as a varieties and does not call into question the same rank for the hispida, he raises the irregularis to the level of an independent species. Further he describes a new species, Keratella earlinae, obviously the North American equivalent to the European K. irregularis, which has not been found in North America so far. K. earlinae as well as K. crassa Ahlstrom seem to be true new world species. They occur widely spread over the continent but have been mistaken without doubt, for K. cochlearis by many authors. (ref. ID; 1402)
ref. ID; 1923
Keratella cochlearis is probably the commonest planktonic rotifer. Carlin (1943) proposed to throw the familiar name K. cochlearis into synonymy with K. stipitata Ehrenberg on the basis of a presumed error by Ehrenberg in drawing a K. quadrata dorsal pattern in a K. cochlearis outline. Some authors have accepted Carlin's suggestion, but there has also been vigorous objection (Berzins 1954), and it may be hoped that K. cochlearis will not be replaced. These animals may be assignable to K. americana Carlin (= gracilenta Ahlstrom), but there are differences in the dorsal sculpture from Ehrenberg's figure. (ref. ID; 1923)
ref. ID; 2393
According to Ahlstrom (1943), Lauterborn's five series mentioned above are to be as follows:
1. The macracantha series belongs to the typical K. cochlearis. In the specimens of the United Sates, the median line on the dorsum has a slight jog on the right-side at a point between the first and the second plaques, while the median line is straight in most cases of European specimens. Both types are also found in Japan.
2. A new combination of names was proposed by Ahlstrom to the typical form of the irregulars series which should be a species separate from the present species. This is seldom seen in Japan.
3. The robusta series is a large-sized group, in which the median line does not jog. This type is rare in Japan.
4. The variety hispida, too, is larger size; the whole surface of lorica is hispid, and sometimes the fundamental patterns of dorsum are indistinct. This is rarely met with in Japan.
5. The forma leptacantha Lauterborn was considered by Ahlstrom to be a local variant of the typical K. cochlearis.
Keratella cochlearis is one of the commonest rotifers in Japan. The size of body proper varies considerably, the length of hind spine deviating from nothing to as long as or more than the body proper. It will be realized that variation seem to be caused not only by the conditions of localities but also by the seasonal factors. Generally speaking longspined individuals occur in early season and the progress of generations through repeated parthenogenesis makes the reduction of the length of hind spine. The cyclomorphosis may, however, not occur due to the conditions of habitat water; an acyclic generation is resulted. A good samples of such as acycly due to a large body of water is seen in K. cochlearis in the southern basin of Lake Biwa-ko. The length of hind spines and the hind spine indices do not vary in accordance with the climates. In North America, a small-sized K. cochlearis is said to occur in weakly acid water. On the contrary, a remarkably large type of K. cochlearis is found in the waters of similar condition in Japan, e.g., ponds among the Sphagnum-moors on the Shiga Heights and on Mt. Hakkoda. The size of individuals from the Shiga Heights exceeds that of var. robusta, but they belong without doubt morphologically to the macracantha series. Moreover, it is a particular interest that the length of the hind spine is considerably longer than the body proper. It would, therefore, be called as the megacantha type in the macracantha series. The measurements made on more than one thousand individuals of this series revealed us the following facts worthy to be described. A gap is clearly observed between the megacantha type and the macracantha type, and also between the macracantha type and the tecta type, in the latter of which the hind spine is entirely lacking. On the other hand, individuals of intermediate types are continuous in regard to the features which are measured, either the length of body proper and hind spine or the hind spine indices. Anyhow, there can be seen a tendency that the larger the body size, the longer the hind spine. (ref. ID; 2393)
ref. ID; 2553
For reasons of priority Carlin (1943, p.55) is of the opinion that this species ought to be called Keratella stipitata Ehrenberg. Berzins (1954) does, however, not approve Carlin's motivation, and the present author agrees in this respect with Berzins. In view of the different opinions expressed the author has referred the question, in accordance with the decision of the Zoological Congress in Paris of 1948, to the International Commission on Zoological Nomenclature (see Bull.Zool.Nomencl.4, p.76, 1950). Pending the decision of the Committee the author uses the, up to now, most current name, viz. Keratella cochlearis. (ref. ID; 2553)
ref. ID; 3226
The Keratella species with one posterior spine and size anterior spines, and a marked median keel was, without doubt, Keratella cochlearis (Gosse). It is difficult, however, to classify the form within the 8 varieties and 6 forms described (Voigt 1967: 177-180). The identification of varieties and forms is based on the following characteristics: total length of posterior spine, macro- and micro-sculpture dorsally on lorica, thickness and colour of lorica and the angle between posterior spine and ventral list of the lorica. Different varieties may show cyclomorphosis. [Lake Blankvatn specimens] The lorica was thick and yellowish brown. The angle between the posterior spine and the ventral list of lorica varied. The total length and the length of the posterior spine were measured on 40 individuals at different seasons. The individuals tended to be larger in spring that in summer and winter. The proportion between total length and length of posterior spine nearly always seemed to be a constant 10:3. Individuals without posterior spine were not observed. The ornamentation of the lorica was not quite uniform. The lorica had two or three lateral plaques. Post-carinal plaques were more less fully developed. The median keel was either straight or slightly curved into the first left carinal plaque. It was difficult to decide whether the micro-sculpture was large or small. Judging from drawings by Latuerborn (1900 taffel 1), it would be median sized. Dorsally there were no spines on the lorica. After careful study of the works of Ahlstrom (1943), Voigt (1957: 177-180), Pejler (1962) and Sudzuki (1964), the form in Blankvatn was found to bear closest resembles to Keratella cochlearis var. robusta (Lauterborn). In lakes with low production, however, Pejler (1962) found that K. cochlearis had only a very slight cyclomorphosis and the macro sculpture of many individuals from Blankvatn was much like Lauterborn's drawings of K. cochlearis cochlearis. Pejler (1962) fond great morphological differences with the group robusta and suggested that several sibling species might be contained into this group. (ref. ID; 3226)
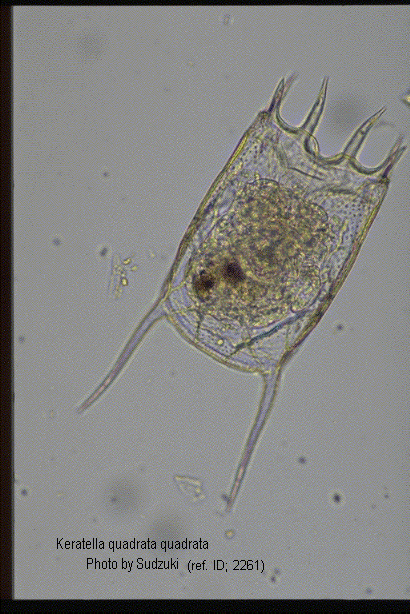
Keratella quadrata (O.F. Muller, 1786) and K. valga (Ehrenberg, 1834) were placed together with several other varieties under a single species Anuraea aculeata Ehrenberg, 1832. It is because the foundation patterns on the dorsum as well as the position and the length of the posterior spines are quite uniform in them. After examining extensive materials from South Africa and India, Edmondson & Hutchinson (1934) were fully convinced of the correctness of the contention that K. quadrata and K. valga are to be recognized as two distinct species. The correct designation for the species was proposed for the first time by these authors. According to Edmondson & Hutchinson (1934), K. quadrata has the following characters. 1. maximum breadth of lorica slightly greater than the posterior breadth. 2. posterior portion is greater than the anterior breadth. 3. two posterior spines are usually subsequal. According to Edmondson & Hutchinson (1934) this species comprises the following nine formae, f. platei, f. divergens, f. frenzeli, f, quadrata, f. testudo, f. brevispina, f. valgoides, f. irregularis and f. curvicornis. Later Carlin (1943) divided this species precisely into the following four 'groups', viz, frenzeli (with var. platei,) quadrata, dispersa and reticulata, while Ahlstrom (1943) into six 'varieties', viz, adnata, platei, edmondsoni, brevispina, curvicornis and pyriformis besides the typical species quadrata quadrata (with formae, testudo, araneosa). Gillard (1948) treated this species as K. quadrata OE quadrata, K. quadrata OE frenzeli respectively, and Voigt (1957) adopted Carlin's system adding one form klementi. Thus, the treatment of formae, and variety of this species still contains some unresolved problems. (ref. ID; 3114)
ref. ID; 3513
Keratella quadrata (Muller) and Keratella valga (Ehrenberg): In spite of the valuable contributions of Hartmann (1918) and others, there still appears to be considerable uncertainly as to the status and correct designation of the various forms included by Harring (1913) under K. quadrata (Muller). After examining extensive material from South Africa and India we are fully convinced of the correctness of the contention that two species, K. quadrata (= aculeata Ehrenberg) and K. valga, are to be recognized. Moreover, the nomenclature of these two species in their typical forms raises no difficulties. Anuraea aculeata Ehrenberg (1832) is expressly stated by its describer (1832) to be identical with Brachionus quadratus Muller. Ehrenberg's best figure (1838, T.LXII, fig.xiv, 1) may therefore be taken in conjunction with those of Muller (1786, T.XLIX, figs.12, 13) as defining the typical form of quadrata. The posterior margin of the lorica in Ehrenberg's figure, measured across the outer margins of the bases of the posterior spines, is wider than the anterior margin similarly measured across the bases of the antero-lateral spines. The posterior spines are parallel (Muller) or very slightly divergent, about two-fifths as long (Ehrenberg) or half as long (Muller) as the lorica. Both reticular and punctate sculpture are clearly indicated by Ehrenberg. Muller's figure is so unsatisfactory that in fixing the typical form it is best to abide by the excellent illustration given by Ehrenberg, save that it is convenient to regard typical quadrata as having practically parallel posterior spines, as is indicated by Muller. Anuraea valga Ehrenberg (1834) may be regarded as typically represented by Ehrenberg's best figure (1838, T.LXII, fig.xv, 1), which shows a form in which the posterior width of the lorica is distinctly narrower than the anterior. The right spine is about one-third, the left one-quarter of the length of the lorica. Reticulate and punctate sculpture are both clearly indicated. In examining as much material and as many illustration as are available to us we have rarely had any difficulty in assigning individuals or illustrations to one or the other species. It is true that in a few figures of European specimens (e.g. Virieux 1916, fig.46; Wesenberg-Lund 1930, Plate VII, figs.8, 26, and 46) the anterior and posterior borders are equal or the former very slightly longer than the latter. But such specimens do not appear to show the pronounced narrowing of valga and it must be remembered that a very minute error in drawing on the part of an observer not studying the dimensions in question would produce just such differences as found in some of these figures. Fedeev (1927) used the relation between the anterior and posterior widths of the lorica to separate tropica Apstein from the other forms of K. quadrata (s. lat.), apparently understanding by Apstein's name what we here regard as K. valga. Within these species the use of misapplied varietal terms appears to have led to considerable confusion. Thus in his monumental work on the cyclical phenomena in rotifers Wesenberg-Lund (1930) objects to certain of Hartmann's conclusions as to the production of valga forms from resting eggs. But it is clear from a study of Wesenberg-Lund's figures that what this author calls valga is a form of quadrata (s. s.) while Hartmann's observations on this point referred primarily to the true K. valga (Ehrenberg). Since such confusion is bound to result from the present unsatisfactory state of the varietal nomenclature of these two species we have attempted to standardise as far as possible the names of the various forms, introducing a minimum of new names and adhering to the principle of priority. Though the latter is not binding in such cases, it would appear to provide the best method of determining the relative merits of two synonym, and the neglect of the principle in the past has led authors to create new varietal name without an adequate study of the literature so that the form of valga with a single posterior spine has been provided with at least thee and probably four names. Kratzschmar (1908, 1913), Hartmann (1918) and Wesenberg-Lund (1930) have studied the morphological cycle in this species. Normally the first generations from sexual eggs are composed of forms with long divergent spines (f. divergens Voigt); these later give place to forms with shorter more parallel spines (f. quadrata Muller). Later one (f. valgoides n.) or both spines (f. curvicornis Ehrenberg) are lost and at such periods sexual reproduction is stated to occur. The amount of cyclomorphosis is very variable in different localities but in general this species exhibits a cycle of reduction. In a few cases an initial elongation of the spines has been observed and in a series called by Hartmann "A. aculeata-valga", but consisting of apparently morphologically normal reduced forms of K. quadrata, the cycle begins with curvicornis, to which the addition of one minute papilliform spines produces the form named by Jakubski (1915) irregularis, and two minute spines testudo Ehrenberg. In general the two posterior spines of quadrata are subequal in length, but in certain forms unequal spines apparently indicate a transition from divergens (right spine) to quadrata (left spine). Fadeev has figured as valga a specimen of quadrata with unequal subparallel spines but this may best be regarded as a transition to valgoides, though Wesenberg-Lund's figures and our collection from Tso Nyak indicate that the latter form may probably appear in a quadrata population without intermediate forms with unequal spines. The following names are probably sufficient to describe the chief members of the cycle and to designate the varieties present in single collections. (ref. ID; 3513)
Keratella tropica var. aspina Fadeew, 1927 (ref. ID; 3688)
Syn; Keratella quadrata var. tropica f. aspina Fadeew, 1927 (ref. ID; 3688)
Keratella valga-group
ref. ID; 3114
Keratella quadrata (O.F. Muller, 1786) and K. valga (Ehrenberg, 1834) were placed together with several other varieties under a single species Anuraea aculeata Ehrenberg, 1832. It is because the foundation patterns on the dorsum as well as the position and the length of the posterior spines are quite uniform in them. After examining extensive materials from South Africa and India, Edmondson & Hutchinson (1934) were fully convinced of the correctness of the contention that K. quadrata and K. valga are to be recognized as two distinct species. The correct designation for the species was proposed for the first time by these authors. Under this species Edmondson & Hutchinson (1934, p.170-171) distinguished eight formae based upon the condition of the posterior spines. They are: f. valga, f. asymmetrica, f. monstrosa, f. longicornis, f. tropica, f. reducta, f. brehmi and f. aspina. Carlin divided these formae into three, namely valga, tropica and procurva. And Ahlstrom (1943, pp.448-453), var. procurva, f. typica, f. brehmi respectively. Recently, Berzins (1955) reviewed thoroughly this valga complex, and distinguished four species and one subspecies, namely K. valga, K. tropica, K. procurva (f. valga after Edmondson & Hutchinson), K. lenzi and K. lenzi var. heliaca, all based upon the posterior area of the dorsum. However, the question still remains unanswered, since Edmondson & Hutchinson simply divided the specimens on the basis of the length of the posterior spine, while, Berzins did it on the basis of the dorsal pattern of the lorica. (ref. ID; 3114)
Keratella valga (Ehrenberg, 1834) (ref. ID; 1345, 1350, 1803, 2277, 2393, 2757, 2816, 2836, 3031, 3114, 3264, 3688) reported year? (ref. ID; 3029, 3065, 3126, 3208, 3484, 3513, 5022, 5031) reported author and year? (ref. ID; 2871, 5389), valga valga (Ehrenberg, 1834) (ref. ID; 1345, 2860, 3073, 3083), valga f. valga Ehrenberg (ref. ID; 3523)
Quote from ref. ID; 3073
Syn; Anourella valga Dujardin, 1841 (ref. ID; 3688); Anuraea aculeata valga Skorikov, 1896 (ref. ID; 3031) or Skorikow, 1896 (ref. ID; 3688); Anuraea aculeata valga Weber, 1898 (ref. ID; 3688), var. valga Weber, 1898 (ref. ID; 1345, 3031); Anuraea procurva Thorpe, 1891 (ref. ID; 1345, 2836); Anuraea scutata Thorpe, 1891 (ref. ID; 1345, 2836); Anuraea valga Ehrenberg, 1834 (ref. ID; 1350, 2277, 2757, 2836, 3031, 3126, 3208, 3688); Anuraea valga assymetrica Barrois & Daday, 1894 (ref. ID; 3688); Keratella aculeata valga Rodewald, 1935 (ref. ID; 3688); Keratella quadrata Edmondson & Hutchinson, 1934 (ref. ID; 3031); Keratella quadrata Harring, 1913 (ref. ID; 3688); Keratella quadrata valga Fadeev, 1927 (ref. ID; 3031) or Jakubski, 1921 (ref. ID; 3688); Keratella valga Carlin, 1943 (ref. ID; 3031)
Keratella valga f. aspina Fadeew (ref. ID; 1345) or Fadeev (ref. ID; 3513) reported author and year? (ref. ID; 3114)
Keratella valga f. asymmetrica Barrois & Daday, 1894 (ref. ID; 1350) reported year? (ref. ID; 3513) reported author and year? (ref. ID; 3114), var. asymmetrica (Barrois & Daday) (ref. ID; 1345, 3523)
Keratella valga f. brehmi Klausener, 1908 (ref. ID; 3688) reported year? (ref. ID; 1345, 3031, 3513) reported author and year? (ref. ID; 3114)
Syn; Anuraea curvicornis Klausener, 1908 (ref. ID; 3688); Anuraea curvicornis f. brehmi Klausener, 1908 (ref. ID; 3031)
Keratella valga f. heterospina Klausener, 1908 (ref. ID; 3688)
Keratella valga f. longicornis Schmarda (ref. ID; 3513) reported author and year? (ref. ID; 3114)
Keratella valga f. monospina Klausener, 1908 (ref. ID; 3688)
Keratella valga f. monstrosa Barrois & Daday, 1894 (ref. ID; 1350) reported year? (ref. ID; 3063, 3513) reported author and year? (ref. ID; 3114), var. monstrosa (Barrois & Daday) (ref. ID; 1345)
Keratella valga f. reducta (Fadeew) (ref. ID; 1345) or Fadeev (ref. ID; 3513) reported author and year? (ref. ID; 3114)
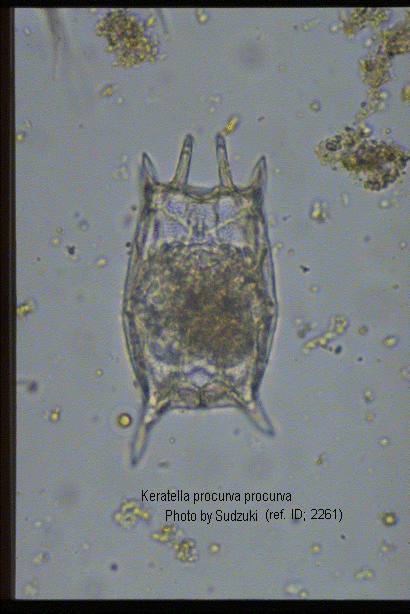
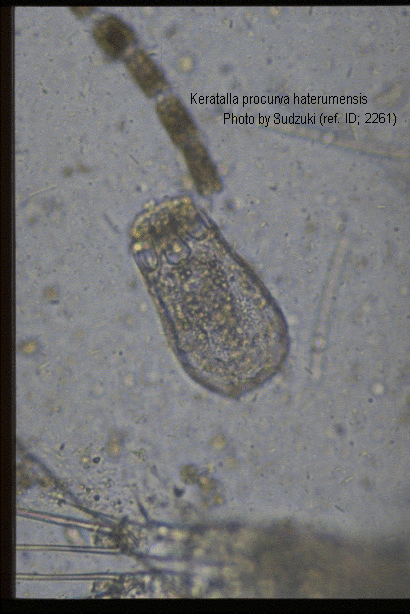
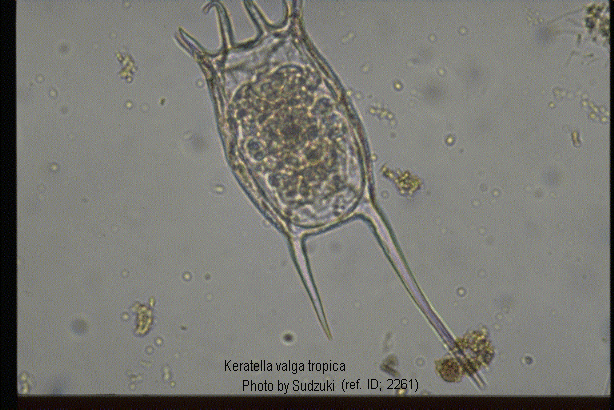
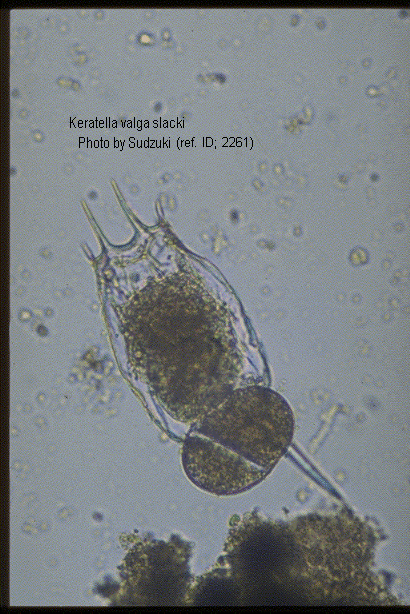
Keratella valga tropica (Apstein, 1907) (ref. ID; 2891, 3083) reported year? (ref. ID; 3062) reported author and year? (ref. ID; 3073), f. tropica (Apstein, 1907) (ref. ID; 1345, 1350, 2064, 2261) reported year? (ref. ID; 3063, 3513, 3523) reported author and year? (ref. ID; 3114, 3292), var. tropica Apstein (ref. ID; 3262)
Quote from ref. ID; 2261
Keratella valga f. tropica-asymmetrica Apstein-Barrios & Daday (ref. ID; 3484), var. tropica-asymmetrica (ref. ID; 1350)
Keratella valga f. tropica-monstrosa (Barrois & Daday) (ref. ID; 3484)
The lorica is tub shaped and moderately compressed dorso-laterally. The greatest width is at approximately three-quarters of the body length, and is five-ninths of the length. The depth is two-ninths of the length. There are six anterior spines the medians being the longest, slightly of converging, and bent ventrally at approximately 90 degrees. The intermediate spines are straight slightly inclined ventrally, and almost the same length as laterals. The laterals are recurved. Two very small posterior spines are directed ventrally. The dorsal pattern is a modification of that of K. sancta Russell, 1944. The anterior plaque is a quadrilateral the posterior side often being indistinct; posterior to this is a triangle with its apex pointing backwards and near the median. Posterior to this on the left is an acute diagonal triangle, and on the right an approximate five-sided figure. Posterior to the acute triangle is another approximate quadrilateral. In the rear of this are two open plaques separated by a Y division. Both posterior plaques contain small triangles. The pattern is that of K. sancta on which has been superimposed an acute diagonal triangle, and a diagonal like commencing from the right anterior side, and extending to the left posterior side. The dorsal plate is heavily pustulated, the pustulations being heavier round the edges than in the central plaques. (ref. ID; 3126)
Comments
K. ahlstromi differs from K. sancta in two respects: the recurved anterior lateral spines, and the dorsal pattern which is constant amongst specimens. In addition K. sancta has so far only been found in highly alkaline fresh waters, and in brackish habitats, whereas the new species was taken in acid water where it is from not common to rare. (ref. ID; 3126)
Etymology
This species is named in honour of Dr. A.H. Ahlstrom of Stanford University, California. (ref. ID; 3126)
Measurements
Length overall 190-195; maximum width 100-110; depth 40-45; spines med. 35-45, int. 20-25. Lat. 22-27 µm (n=5). (ref. ID; 3126)
The lorica is not as slender as shown by Ahlstrom (1940) and resembles K. cochlearis, except for the pattern of the lorica where a series of median plates can be recognized. The animal is smooth like those described by Hauer (1952) from Lake Maracaibo in Venezuela, not covered with pustules (cf. Thomasson 1953). (ref. ID; 2894)
Relatively broad and thickset dorsal shell with a large median frontal field which is enclosed on both sides by a small lateral frontal field. An oblong anteromedian field is to be seen. The underlying facets, a mesomedian and a posteromedian field, are not separated by a border. The latter and a pair of postero-carinal-facets are enclosed by a marginal field. The caudal-spine inserts displaced to the right. A similar insertion shows K. mixta (Oparina-Charitonova, 1924) belongs to the cochlearis-forms because of the configuration of the dorsal fields. Lateral marginal fields in the upper lorica are not to be seen in top view. The anterior ventral margin of the shell and the ventral plate are built normally. (ref. ID; 2834)
The highly recurved form of this stoutly built, narrow-bodied Keratella is unlike any known species. The reddish-brown lorica is widest at the tips of the anterolateral spines (71 µm) and narrows (43 µm) just below the base of the anterior spines to form a distinct neck-like region. The midbody region widens again (49 µm), giving an undulating curvature in dorsal view. The posterior spine (~111 µm) is longer than the body (~100 µm), composing ~1/2 of the total lorica length (~218 µm). Posterior spine is somewhat dorsoventrally flattened near the base but becomes more rounded posteriorly. The posterior spine forms an angle with the long axis of the body which varies from about 40 to 75 degrees. Ventral and dorsal plates are both arched so that the body has a circular or ovoid cross section. The anterolateral spines (25.5 µm) are only slightly shorter than the anteromedial ones (29.4 µm) and are much longer than the anterointermediate spines (9 µm). The anterior spines and the anterior margin of the ventral plate form a cage-like opening. A thickened ridge transverses the dorsal plate just below the base of the anterior spines. The faintly developed facet pattern of the dorsal plate is similar to that of K. cochlearis and K. taurocephala. The medial frontal facet and dorsomedial ridge are faintly visible with light microscopy and are clearly seen with the scanning electron microscope. The ridge veers diagonally to the right near the middle of the body for a short distance before continuing posteriorly to the base of the spine. This feature is prevalent on North American taxa belonging to the K. cochlearis group of morphologically similar species. At the angles of this bend a lateral ridge runs perpendicular to the main axis of the body where it bifurcates to form the anterocarinal facets. The reticulate network of the surface of the lorica is composed of pentagonal and hexagonal elements which lack spinelets on both dorsal and ventral surfaces. The lateral antennae are located near the midbody region and not in the lower 1/2 of the body, as is characteristic of other species in the genus. The ventral plate is well-developed anteriorly but seems to thin posteriorly, having prominent longitudinal folds near the vicinity of the cloacal opening. Two prominent papilla-like structures within the body cavity project into depressions on the inner surface of the dorsal plate near the middle of the lower 1/3 of the body. Pores (1 µm diameter) connecting these gland-like sacs arise on the dorsal lorica surface and are located ~10 µm in each side of the mid-dorsal ridge directly above the sacs. These structures have not been reported in other species of Keratella and may have a secretory function, possibly producing mucus for temporary attachment. (ref. ID; 7256)
Male: Males of Keratella armadura were not observed and are unknown in the species. (ref. ID; 7256)
Notes
Keratella armadura superficially resembles K. taurocephala. The lorica of the former is stained a reddish brown whereas that of K. taurocephala is translucent and usually covered with spinelets on both plates. The anterolateral spines are also not as larger or laterally divergent as those of K. taurocephala. The body of K. armadura is much narrower and more rounded in cross-sectional view than that of K. taurocephala, and the posterior spine is considerably longer and more acutely bent. (ref. ID; 7256)
Etymology
From Latin; the specific name armadura ("hard armor") refers to this species' rigid lorica and hard spines. (ref. ID; 7256)
Type material
Holotype: National Museum of Natural Sciences, Ottawa, NMCI1989-0123. Single female mounted on a glass slide in glycerine with 5% buffered formalin and sealed with Permount. Individual taken from a population collected from Mud Lake Bog (45 degrees 36'30"N, 84 degrees 36'00"W), Cheboygan Co., Michigan, on June 10, 1974. (ref. ID; 7256)
Paratypes: National Museum of Natural Sciences, Ottawa, NMCI1980-0124. Thirteen females taken from the same collection as the holotype also mounted on a glass slide in glcerine with 5% buffered formalin and sealed with Permount. (ref. ID; 7256)
The narrow occipital region, the broad posterior region, and the arched posterior spines are shown in figure. The dorsal pattern and tiny but prominent postulations are clear. The foundation pattern is typical of K. quadrata, though not very sharp. The occipital ventral margin is not clear but the anal opening is shown figure. (ref. ID; 2598)
Comments
According to Ahlstrom the important criteria for separating most varieties and forms in K. quadrata depend on the foundation pattern of the dorsum which has to be used in conjunction with other features, including posterior spines. In the drawings showing the "quadrata" group, he has given the outline of a form of K. quadrata without any dorsal pattern from material collected from Clark Lake, Saskatchewan, and he says "sculpture indistinct". Berzins obtained some material from Southern Indian Lake, Manitoba, too, and found a form similar to the one described by Ahlstrom. Berzins observed the foundation pattern in the occipital region and posterior part of the dorsal plate. The nature of the foundation pattern, the peculiar nature of the posterior spines, and the narrowness of the body at the occipital region warranted Berzins to create a new species Keratella canadensis. (ref. ID; 2598)
Measurements
Total length 331-382; width 105-128; width occipitally 78-92; width posteriorly 125-140; length of body 162-178; length of posterior spine 132-167; occipital spines, lateral 38-46; occipital spines, intermedian 45-51; occipital spines, median 54-74 µm. (ref. ID; 2598)
Keratella carinata Russell, 1950 (ref. ID; 1923, 3181 original paper) or 1952 (ref. ID; 2280)
The lorica terminates in a short, stout, recurved spine which is slightly off the median line. The maximum width of the lorica is approximately seven-tenths of its length, and occurs slightly before mid-length. The depth of the lorica is approximately five-elevenths of its length, is almost constant for three-quarters of its length when it falls sharply to the posterior spine. There are six anterior spines. The medians are slightly diverging, and at approximately one-quarter their length are bent ventrally 90 degrees. The length of the median is approximately seven-tenths of the lorica. The intermediate spines are slightly diverging, straight, and inclined to the ventral at 45 degrees. The lateral spines proceed from the anterior lateral edge of the lorica normal to the ventral surface. In a few specimens the lateral were slightly inclined posteriorly. The pattern of the dorsal plate is very irregular. There are two closed anterior plaques, a hexagon and a pentagon, followed by two pentagons, a hexagon and a pentagon, arranged in an approximate semi-circular. Posterior to these are a pentagon, at two hexagons. Two open plaques complete the pattern. The dorsal plate is areolated except in the central area, the markings being minute. The dorsal pattern, although unsymmetrical, is constant between specimens except that the sides of the plaques on the extreme edge to the lorica are sometimes missing. The main features of this species are: the dorsal pattern, the depth and shape of the lorica with its almost vertical sides, and the length, inclination, and shape of the anterior spines. (ref. ID; 3181)
Comments
Although K. carinata differs in the length and shape of the anterior and posterior spines, the dorsal patterns which are of an unusual type are identical, and from a re-examination of the material I have no hesitation in saying that K. carinata must be considered a synonym for K. javana Hauer. (ref. ID; 2280)
K. carinata resembles K. irregularis and K. mixta, but only superficially. It resembles the latter from a dorsal view, but the posterior spine is not so much off the median line and the lateral spines of K. mixta are entirely different. The median and intermediate spines of K. carinata are longer and more acutely bent. The similarity with K. irregularis is confined to the dorsal view, the anterior spines of the latter are different in length and position, and the posterior spine is decurved instead of being recurved. The dorsal pattern of K. carinata is entirely different from that of any other species. As Ahlstrom (1943) has stated, "The foundation pattern of the dorsum is constant in any species, and is a highly important taxonomic character". The internal organs of K. carinata are normal to the genus. (ref. ID; 3181)
Type locality
K. carinata was collected by Professor Pereival from peaty roadside pools at Kumara, Westland. From the nature of the material it is assumed that the water was of acid content. (ref. ID; 3181)
Lorica pattern constant with open posterior carinal plaques, no pustulation. Body oblongata and narrow, spine usually 1/4 of total length. Rather strong temporal variability, longest in June, shortest in August/September. The smallest individuals are to be found in the warmest parts of the lake. A slight allometry appears in the body/spine ratio, the spine being relatively shorter in late summer. (ref. ID; 1402)
Lorica with a stout median posterior spine. Dorsum with characteristic median longitudinal line, with symmetrically arranged plaques on either side. (ref. ID; 1929)
The dorsal pattern was absent and the pustulation very faint. (ref. ID; 3127)
Lorica terminating in a stout median posterior spine which usually varies in length from as long as the body proper to one-third as long but which may be even more reduced or completely wanting. Anterior dorsal margin with six spines; medians longest, curved ventrally; intermediates usually slightly divergent, somewhat shorter than lateral spine which are convergent at their tips and which arise at a single angle toward the ventral. Foundation pattern of the dorsal plate characterized by a median line extending longitudinally from behind the median frontal area to the base of the posterior spine. Only one pair of fully enclosed carinal plaques (antero-carinal plaques) often present. (ref. ID; 3180)
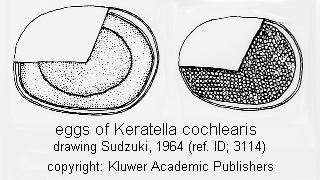
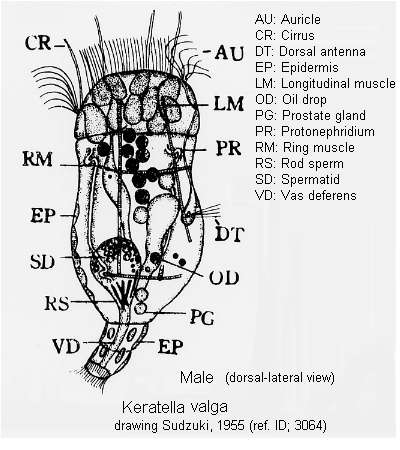
Egg: (ref. ID; 3114)
Quote from ref. ID; 3114
Measurements
Total length 110-180 µm. (ref. ID; 1402)
Total length 154; width of the anterior margin 56; width of the posterior margin 42; length of lorica 84; posterior spine 56 µm. (ref. ID; 1929)
Total length 211; length of body 99, width 67; anterior lateral spine 29; anterior intermediate spine 25; anterior median spines 32; posterior spine 80 µm. (ref. ID; 2715)
Total length 95-156; dorsal lorica length (without spines) 80-90; width at the basis of occipital lateral spines 40-50; widest part 40-60; height at the base of frontal spines 30; highest part 36; anterior median sinus 15-20; anterior intermediate spines 10-12; anterior lateral spines 8-18; caudal projection or protuberance 0-48 µm. (ref. ID; 3083)
Total length 150; length of lorica 80; length of posterior spine 30 µm. (ref. ID; 3127)
Length of lorica 112-120; width of lorica 60-70; length of median occipital spines 16-24; length of lateral occipital spines 12-18; length of posterior spine 25-45 µm. In a few cases we saw individuals with a large egg 55x40 µm. (ref. ID; 3275)
The dorsal pattern is almost entirely obscured by the heavy pustulation. (ref. ID; 2284)
Comments
Pejler's (1962) figures of Keratella cochlearis var. hispida from central Sweden show markedly hispid loricae. However, in the forms found in Ontario the hispidity, though variable in different specimens, was found not to be as thick as in the European forms, as noted by Ahlstrom (1943) for North American material. (ref. ID; 3086)
Measurements
Total length ranges from 130 to 225 µm with a strongly expressed seasonal variation. The largest animals appear in July, the smallest one in September. The main size variation is between 165 to 210 µm. There is a significant allometry in the body length: spine length relationship, the ratio varying from a minimum of 2.7 in June to a maximum of 3.6 in September. No clear relationship between water temperature and appearance and variation of the hispida form can be established on the material. (ref. ID; 1402)
Length of body 80; maximum body width 43; length of posterior spine 26; length of anterior spines (median 28, intermediate 12, lateral 12) µm. (ref. ID; 2284)
The status of Keratella cochlearis irregularis is still open to discussion, and has ranged between "var." and bona species (Kutikowa, 1970). We here formally adopt an intermediate position, noting this form as a sub-species to K. cochlearis. (ref. ID; 2825)
Keratella cochlearis javana Hauer, 1937(ref. ID; 3083), var. javana Hauer, 1937 (ref. ID; 3523 original paper)
See
Keratella javana (ref. ID; 1350, 3258, 3274)
Descriptions
The dorsal ornament of the lorica is just like that of Hauer 1937). The specimen from Singapore is always characterized by having strong serration along median spines. The following are in common with Russell's specimen: 1) both median and lateral spines are bent ventrally at 90 degrees, 2) intermedians are inclined to the ventral at 45 degrees, 3) a straight and short ridge is found on the base of caudal projection. According to Berzins (1953: 6) K. carinata Russell, 1950 is synonymous with this species. In this point, Russell (per. com.) and Sudzuki quite agree with Berzins, since there was recognized neither morphological nor ecological differences between the two. It is rather interesting to mention that 1) population density of this species seems extremely low, e.g. only a single specimen has been collected from Java (Hauer 1937), Western Australia (Berzins 1953) and Malaysia (Fernando et al. 1981), 2) javana is considered to be divided into 2 groups, A and B. Group A: Total length (TL) 125-140, i.e. <150 (smaller); caudal projection or protuberance (CP) 10-20 (shorter); CP/TL 0.07-0.16. To this group belong the specimen from New Zealand (Russell 1950: 162-164, Figs.1-2) and Japan (Yamamoto, 1960: 394-395, Fig.7j). Group B: Total length (TL) 156-189, i.e. >150 (bigger); caudal projection or protuberance (CP) 37-42 (longer); CP/TL 0.21-0.27. To this group belong the specimen from Java (Hauer 1937: 382, Fig.29), Zaire (Gillard 1952: 341, Fig.1) and Malaysia (Fernando et al. 1981: 212, Fig.12). The specimen from Singapore seem to belong to group B. (ref. ID; 3083)
All the specimens observed had long posterior spines and the network (areolae) of the dorsal plate continued along the anterior half of the posterior spine. (ref. ID; 3235)
Measurements
The total length equalled 179-227; breadth 63-90; the two median anterior spines 21-30; posterior spine 60-94 µm. (ref. ID; 3235)
Body shape ovate, with rounded margins. Average spine length approximately 1/3 of total length. Variability is not significant. (ref. ID; 1402)
See description of Keratella cochlearis-group. (ref. ID; 2989)
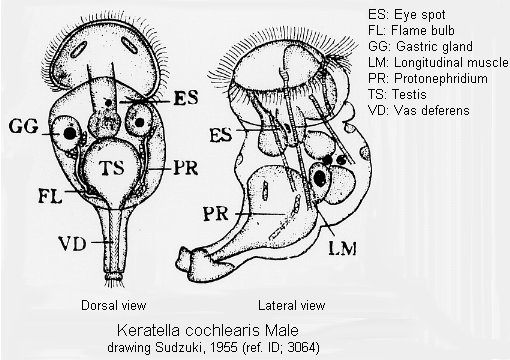
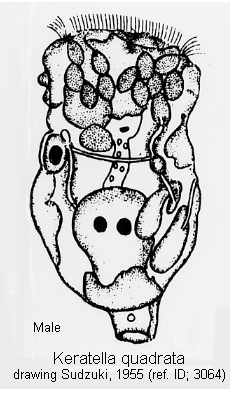
Male: Male present. (ref. ID; 3114)
Quote from ref. ID; 3114
Measurements
Total length, through winter and spring 185-220, average 210. Largest individuals in summer, total length 220-260, average close to 230 µm. (ref. ID; 1402)
One caudal spine. (ref. ID; 1402)
Dorsal plate areolations heavy and no pustulation on either plate. Ahlstrom says that this is a transcursion species being found in acid and alkaline waters, but apparently this is the first time it has been found in a marine habitat. (ref. ID; 2281)
Measurements
Total length 187-229 µm; the average ratio total length/spine length is constant around 3.75. A slight increase in total length can be observed sometimes in smaller specimens. (ref. ID; 1402)
Total length 160 µm; length of body 90 µm; W/L O. 71. Anterior spines med. 24, inter. 13, lat. 15; posterior spine 46 µm. (ref. ID; 2281)
In some cases the basal spicules on the claw were absent. A broad mucous groove on the claw gave the appearance of the claw being double. (ref. ID; 3127)
Measurements
Maximum length 220; maximum width 112; length of toe 90 µm. (ref. ID; 3127)
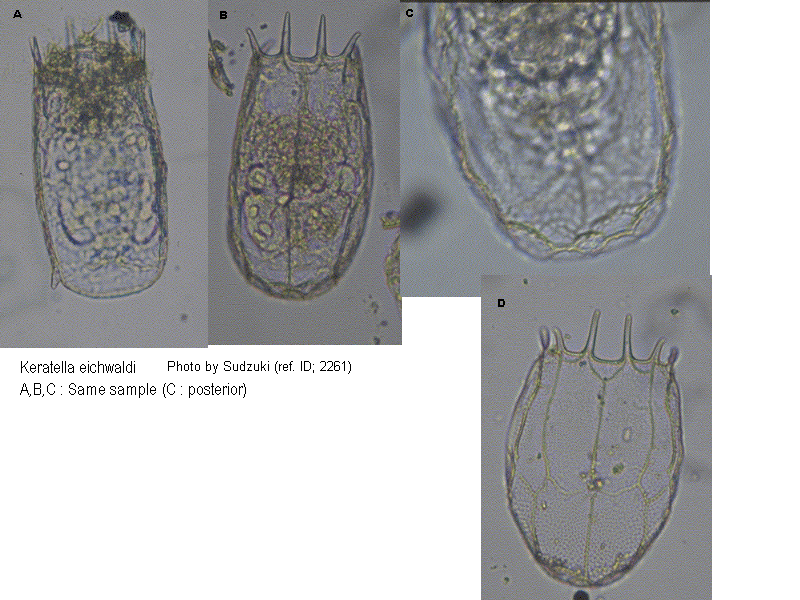
K. cruciformis was described by Thompson (1892) from the sea off the Norwegian coast. K. eichwaldi was described by Levander (1894) from the Finnish coastal waters. The distribution areas of the two species meet in the Baltic and in the Black Sea, at the Donau mouth. The specific differentiation of K. cruciformis and K. eichwaldi is a provisional solution, pending later investigations on variation in transitional areas. (ref. ID; 1866)
Descriptions
This species found chiefly in the sea. Very broad body shape (L/B=1/1). Lorica areolated, two horizontal crests on the lorica. (ref. ID; 1866)
Marine and brackish water. (ref. ID; 3573)
This is brackish-water species. The hind process is absent. Both the punctate and reticular sculptures are remarkable. In the dorsal lorica, the frontal plate is very short (ca. 10 µm), but the first carinal plate is remarkably long (ca. 75 µm). (ref. ID; 3063)
There is a considerable variation in the lorica design, especially in summer. By a reduction of the division lines of the posterior-carinal plaques and the division lines of the median frontal area, the dorsal plaque pattern may change considerably. Further reduction of the lorica design appears at all stages by dissolution of the division lines between the plaques to thinly dotted lines, with increasing pustulation spreading from the anterior parts of the lorica over its entire surface until the form shows similarity with K. cochlearis var. hispida. (ref. ID; 1402)
The lorica has pustules distributed along the ridges of the dorsal plate and also irregularly inside the plaques. There seems to be an extra anterolateral polygon on either side situated between the median frontal area and the accessory anterio-median hexagon which is small. The posterior spine is relatively long. (ref. ID; 2715)
This animal, apparently an American endemity, does not seem to be rare. Ahlstrom described it from Western Canada as well as from Wisconsin, Rawson reported it from Great Slave Lake. I found it in numerous localities: the Georgian Bay of Lake Huron, in the lakes of Mont Tremblant Part (Quebec), lake Memphremagog (Quebec Vermont) and in the St. Lawrence river. The very robust animal can be easily recognized by the peculiar pattern of the lorica. The first median plate is flanked by two carinal plates, which are also connected by a second median plate of the shape of an irregular hexagon. These are followed by two large asymmetrical second carinal plates which are divided by a row of pustules. Finally, these is a third row of carinal plates. The six lateral plates are also irregular in shape. The whole animal is covered with very small pustules. (ref. ID; 2894)
Remarks
The dorsal, medial line of K. cochlearis was unbroken, whereas in K. earlinae it was broken by two accessory medial plaques. This difference was visible throughout the year. Both species showed great variability in pustulation ranging from no to few pustules to many pustules. The size of pustules also varied; some individuals had pustules visible only at x1000, whereas others had pustules visible at x40. Pustulation was generally more obivous in K. earlinae, which also appeared more robust than K. cochlearis. During the study period, both species exhibited temporal variation. For both species, average spine length increased in spine, decreased in summer, and again increased in fall. In winter, average spine-length values appeared lower than in fall. Avergae lorica lengths decreased in summer and increased in fall but appeared stable during spring and winter. The degree of change in spine length of K. cochlearis was much greater than that of K. earlinae. This pattern of temporal variation is considerably different from the usual pattern of long spines in winter and short spines in summer (Lindstrom and Pejler 1975). Lindstrom and Pejler (1975) showed that increased temperature caused shorter spine length in K. cochelaris; however, the spine length of K. cochlearis and K. earlinae in Hastings Lake increased during spring 1975 and 1976 at the water temperature increased from aproximately 10 degrees C in late May to 18 degrees C in late June. Therefore it appears that some factor other than temperature is controlling the variation. It is also possible, however, that similar to King's (1977) study of Asplanchna girodi, each species population may represent a set of genetic subpopulations that dominate the population at different times. Correlation coefficients between lorica lengths of the two species were highly significant (P <0.01) at station A (r=0.07) and C (r=0.81). Coefficients between spine lengths were highly significant (r=0.65, P <0.01) at station A and significant (r=0.45, P <0.05) at station C. Despite the significant correlations, sign test (Sokal and Rohlf 1969) indicated both lengths were greater (P <0.01) in K. earlinae than in K. cochlearis at both station. In fact, the average spine and lorica length were always greater in K. earlinae than in K. cochlearis. On a few sampling dates in spring, however, the extreme values for both measurements overlapped. (ref. ID; 5946)
Measurements
Strong size variability ranging from 120 to 240 with main variations between 175 and 215 total length. Back spine shows only weak allometric variation and is on an average about 1/2 of total length. Spineless forms have been noted but their classification is difficult. It is remarkable that, in spine of a less pronounced average size variation, the smallest individuals found were smaller than the smallest hispida. It is also worth mentioning that the smallest individuals were considerably smaller than given in Ahlstrom's survey (1943). (ref. ID; 1402)
Total length 256; length of body 108, width 89; antero-lateral spine 29; anterior intermediate spine 16; anterior median spines 38; posterior spine 106 µm. (ref. ID; 2715)
Total length 225-235; width 70-75; caudal spine 86-100; frontal spines 25, 19 and 26 µm. (ref. ID; 2894)
This is a doubtful species, because though the median frontal area is similar of that of earlinae, the arrangement of the carinal plaques is quite different and is related to that of K. cochlearis robusta. Sudzuki cannot regard it as a variety of K. earlinae. (ref. ID; 3114)
K. edmondsoni differ from K. quadrata in the absence of posterior-most dorsal median plaque. (ref. ID; 1803)
The arrangement of facets on the dorsal lorica is similar to that of K. procurva (Thorpe). The species differs from K. procurva by 1) the terminal end of the dorsal ridge being bifid anterior to the posterior spines in K. edmondsoni, smoothly ending in K. procurva and 2) posterior marginal plates are absent in K. edmondsoni, present in K. procurva. Additionally, the general shape of the lorica, greater posterior spine length and less stiff and smooth lorica of K. edmondsoni distinguish the species from K. procurva. K. edmondsoni was originally described as K. quadrata var. edmondsoni from Madras, India (Ahlstrom 1943). However, Nayar (1965) considered that it was sufficiently different from K. quadrata to be considered as K. edmondsoni. (ref. ID; 1831)
This species found chiefly in brackish water. Long, narrow body shape (L/B=1.3/1). Lorica crystal-clear, without sculptures, one crest (mostly none) on the lorica. (ref. ID; 1866)
Nauwerck (1978) described considerably smaller species from Lake Ontario than the original species described by Carlin; Total length was 130-190 µm, the longer posterior spine varying 15-50 µm. There is no doubt that the species is identical at least with some of the specimens recorded by Ahlstrom (1943) as K. quadrata var. brevispina (Gosse), to which it also is closer in dimensions than to the European form. One other difference to Carlin's description, the occasional separation of the anterior lateral plaques, has later been noted for European material as well, e.g. by Pejler (1957). A separation of the two seems not to be justified. As both Ahlstrom and Carlin have published their observations simultaneously, the question of priority may be delicate. However, to him it seems more logical and more practical to follow Carlin's terminology. (ref. ID; 1402)
This species distinguished from Keratella quadrata by its low morphological variability and its occurrence at only low temperatures. Problems involving K. hiemalis, K. quadrata, and K. testudo can also be found in North America. In the same year that Carlin described K. hiemalis, Ahlstrom (1943) published his paper on the revision of the Keratella of North America. Unaware of Carlin's work, Ahlstrom, combined K. hiemalis and K. testudo in to the taxon K. quadrata var. brevispina (Gosse). The species does not have the triangular median facet of K. hiemalis; it is clearly K. testudo based on shape, size, and facet pattern. This Synonymy was correctly recognizes by Koste (1978) and Stemberger (1979). In the same paper, Ahlstrom also described a new forma which he called K. quadrata var. brevispina f. gossei. This form is K. hiemalis without posterolateral spines. These spineless forms occur infrequently in stratified lakes on North America and may co-occur with spined forms. To further obfuscate the taxonomy of these two taxa, Ahlstrom (1943) described a large Keratella (180-356 µm total length) which he called K. quadrata f. testudo. The specimens of this taxon are simply shorter-spined forms of K. quadrata. Thus, greater variation in posterolateral spine lengths occurs in K. quadrata than is typically attributed to the species. Agnes Ruttner-Kolisko do not know if Ahlstrom interpreted these forms as K. testudo (Ehrenberg). Given this history, it is clear that K. hiemalis is more variable, at least in North America, than what Carlin originally described in Europe. Most records of these species in North America have been lumped into K. quadrata, the most frequently cited taxon (Stemberger 1979; Stemberger & Gilbert 1987). In fact, no documented records of K. testudo have been reported in North America until recently (Stemberger 1979). This species occurred in about 15% of the lakes surveyed in northern lower Michigan. K. quadrata and K. hiemalis occurred in 60% and 83% of lakes, respectively. K. hiemalis was a dominant rotifer in most lakes during winter and early spring, which is consistent with its cold-water preference (Stemberger 1990). (ref. ID; 1705)
Nogrady (1988) described giant form from Lake Ontario; total length was 300 µm, caudal spines 98 µm. The normal specimens in the same population measured 185 µm mean total length with a 35 µm caudal spine. The giant form resembled a K. quadrata frenzeli, but had the unmistakable delicate and transparent lorica and lorica pattern of K. hiemalis. Carlin (1943) separated K. hiemalis from K. quadrata on the basis of size, lorica pattern and cold stenotherm temperature preference. It seems that size has to be treated with caution. (ref. ID; 1850)
In some specimens there was no ridge separating the first median carinal plaque from the median frontal area, but both of them were connected by a short vertical ridge. The first median carinal plaque therefore appeared more or less triangular. (ref. ID; 3086)
Descriptions
The species is clearly characterized by its low morphological variability and its occurrence at low temperatures. Posterior spines are straight, about half the length of body, forming a prolongation of its lateral borders. The characteristic pattern of the dorsal plate of the lorica is the triangular, first median facet and the unforked posterior ridges. Both K. hiemalis and K. testudo are distinguished from K. quadrata, which has a larger body size and long, curved posterolateral spines. In all three species the anterior spines are similar. (ref. ID; 1705)
The species is cold stenothermal. Some specimens, while fully grown, had a completely transparent, soft lorica without any visible pattern. (ref. ID; 2593)
The size (total length 170-180 µm, width 71-77 µm) and macro sculpture of the lorica were accordance with the description given by Carlin (1943). The only difference compared with Carlin's drawings was the list between the two most posterior median plaques. This was scarcely developed in most of the individuals. No cyclomorphosis was registered. (ref. ID; 2989)
This species is closely related to K. earlinae. The largest individuals seen were smaller and relatively broader than in K. earlinae but significant differences could not be established except for the plaque pattern. K. irregularis has smooth loricas at the beginning and at the beginning and at the end of the season but became coarsely granulated in a similar way as K. earlinae in summer. The trend is size variation is the same as in K. earlinae. (ref. ID; 1402)
K. javana was described by Russell (1950) from New Zealand under the synonym K. carinata and the New Zealand specimens were generally larger and had longer anterior spines than those from The Chatham Island. (ref. ID; 3127)
Measurements
Length of body 110; width of body 55; length of anterior spines, median 40, intermediate 22, lateral 14 µm. (ref. ID; 3127)
K. lenzi (f. ayui) f. nov. is very similar to K. lenzi lenzi, both in stereoultrastructure and shape, except for the presence of one or two small postero-lateral spines, a slightly wider body outline (both maximum and minimum), and a shorter posteromedian field. Although anterior spines were measured as projections -in dorsal view- of their actual size, lateral and intermediate spines are consistently uniform in both morphs, while anteriomedian spines were significantly longer in K. lenzi (f. ayui).
Dorsally observed, the lorica has a glossiform (tongued-shaped) outline. From the anterior corners, the margins of the lorica diverge smoothly to reach the maximum width at the midbody region, covering into a rounded posterior. The maximum width of the lorica represents on average 64% of its length (excluding spines). Larger specimens are relatively more slender than smaller ones. Of 3 pairs of occipital spines, laterally observed, the median is the longest (approximately 1.5 times longer than the lateral pair). Median spines are slightly convergent and strongly recurved towards the ventral region. Intermedia spines are scarcely shorter than the lateral. A single posterior spine usually arises from the right posteromarginal field. More rarely, there is a second shorter spine that arises from the left field. Caudal spines are slightly divergent or parallel to the body axis. As in other Keratella of the 'valga' superspecies, the right posterior spine is either exclusive, or the longest. The right caudal spine is on average slightly longer than the anterolateral spines, reaching only up to 27% of the lorica length. There is no correlation between lorica and caudal spine length. Twenty-one polygonal fields are clearly identifiable on the dorsal plate of the lorica: 5 median, 4 lateral pairs, and 4 marginal pairs. The fields are delimited by prominent ridges superimosed on a basic reticulate pattern of minute polygons, in the posterior region of the lorica, whereas a strong granulation prevails in the anterior and central areas of the lorica. The frontomedian field is an open hexagon, with its lateral ridges prolonged into the anteromedian spines. In the same way the ridges bordering the frontolateral and frontomarginal fields, reinforce the structure of the other pairs of occipital spines. The anteromedian and mesomedian fields are clearly visible hexagons of regular to slightly irregular shape, and comparable size. As in K. lenzi lenzi, the posteromedian field is an elongate trapezoid with a rounded caudal margin. Between this field and the margin of the lorica there is a narrow fifth postero-median field, or posteromedian remnant. Laterally observed the caudal region is truncated in adult specimens, owing to the remarkable growth of the posteromedian field. This character is much more pronounced in K. lenzi lenzi than in the new morphotype. There are 4 pairs of lateral fields; frontal, anterior, median and posterior. The lateromedian fields carry the lateral antennae in the marginal angles of their posterior third. The location of this pair of minute pores (2 µm) is distinguishable under light microscope but their organization is only revealed by SEM (x18,000). In K. lenzi (f. ayui) these pores are bordered by 9 papillae forming an oval, bulged ring. Although its value as a taxonomic character has not been explored, these pores are likely to be morphologically stable and promising for the investigation of the phylogenetic relationships among loricate rotifers. Only 4 pairs of marginal fields are visible in the material I have examined: frontal, anterior and two posterior. The median pair is either lacking or inconspicuous. The posteromarginal fields consists of one pair of minute caudal areoles (occasionally subdivided), adjacent to the fifth median field, and one pair of triangular fields laterally placed. The anterior third of the ventral plate is ornamented by minute pustulations extending onto the central region of the lorica. The anterior ventral margin is bordered by a cord-like bulge, and presents a median V-shaped notch, prolonged into a median ridge on the first fourth of the ventral plate. The range of morphological variability of the new morphotype has been depicted. Aside from the presence vs. the absence of posterior spines, a scatterplot of the significantly different morphological traits between both morphos reveals that although they overlap, K. lenzi (f. ayui) can be discered from K. lenzi lenzi on the basis of its lorica width and posteromedian field length. (ref. ID; 1309)
Etymology
The name 'auri' refers to a locality close to the dam site at Salto Grande Reservoirs in Entre Rios Province, Argentina, meaning laurel in Guarani language. (ref. ID; 1309)
Lorica elongated, narrow, lacking posterior spines, with characteristic posteromedian field on the dorsum, and massive, short anterior spines. Facets of dorsal lorica with alveolar network. The posteromedian facet large, broad, tongue-shaped, with longitudinal wrinkles. Ventral lorica plate narrower than dorsal plate. Trophi malletae and symmetrical. (ref. ID; 7143)
Differential diagnosis
Keratella mexicana sp. nov. belongs to the Keratella quadrata group, with which is shares the number of facets and a similar structure of the posterior margin of the dorsal plate of the lorica. However, K. lenzi has a wider anterofrontal facet, its second median facet is a regular hexagon, and its extended trapezoidal posteromedian facet continues in a series of easily identifiable, narrow marginal facets (or posteromedian remnant), which are obscured by the overhanging margin of the dorsum in the new species. K. lenzi also differs from K. mexicana in its body outline and the structure of its anterior spines. The new species also has features in common with K. slacki. According to observations by Berzins (1963) and Koste (1979), the long narrow second median facet and, especially, the outline of the posteromedian facet of K. slacki are shared with K. mexicana. There are obvious differences in the pattern of facets, especially in the structure of the posterior margin and in the general shape of the two species, even when the posterior spines are lacking. K. slacki usually has long posterolateral and anteromedian spines, and a relatively large lorica. K. tropica less close to the new species than K. slacki. In K. tropica, the second median facet is a regular hexagon and the posteromedian facet, although elongated, has straight contours. In this species, at least the posteromedian remnant is recognizable, even in specimens with a hardly discernible foundation pattern (Marinone & Zagarese 1991). (ref. ID; 7143)
Descriptions
Lorica rigid, elongate, narrowed and rounded posteriorly, widest at posterior third, approximately same width of anterior margin. Dorsal plate domed, margins thickened. Median frontal facet small, its lateral arc bent, passing to anteromedian spines. Second median facet short, broad, usually hexagonal. Less conspicuous lateral ridges arising from lateral angles of second and third median facets bifurcate terminally, defining lateromarginal facets in some specimens. Lateral antenna located approximately at central region of lorica, near marginal angles of lateromedian facets. Lorica surface of all circumscribed facets with alveolar network but less granulated than in K. cochlearis Gosse, except posteromedian facet. This facet large, broad, tongue-shaped, with peculiar longitudinal wrinkles, resembling the furrows of Notholca verae Kutikova. Posterior edge overhangs posterior margin of lorica. Three distinct lateral ridges arise from posterior margin of fourth median facet. Anterior margin of dorsal plate with three pairs of relatively short but massive spines, bent inwards and slightly convergent at their tips. Widely separated medina spines, longest, strongly thickened, intermediate spines more pointed, lateral spines long, broadly based, with thin, pointed tips. Ventral lorica plate narrower than dorsal plate, usually slightly concave (in some specimens second half of lorica convex), with smooth surface. Its anterior margin V-shaped, bordered by cord-like bulge. Posterior margin of ventral plate trilobate, distinctly separated from posterior margin of lorica, the latter constricted laterally, having longitudinal wavy wrinkles. Trophi (20 µm long, 28-32 µm wide). Trophi malleate, symmetrical. Uncus bearing six teeth, their size decreasing from first to sixth, first and second fused at their bases. Internal margin of rami (12 µm) with six impressions corresponding to teeth on each uncus. Fulcrum short and narrow. Manubrium rather massive (14 µm) with three cameras of different size and moderately short cauda. Salivary and gastric glands small. Oesophagus short. Large pouch-shaped stomach. Intestine damaged in preserved material. Vitellarium large, eight nuclei. (ref. ID; 7143)
Male: Male unknown. (ref. ID; 7143)
Etymology
The new species' name is an adjective, given after the Republic of Mexico. (ref. ID; 7143)
Type locality
A small reservoir situated at 21 degrees 59'N, 101 degrees 59'W, surface area 4125 m2, altitude 2025 m over sea level, depth ranges from 0.2 to 2 m. It is located at 7 Km W from Palo Alto, Aguascalientes, Mexico. (ref. ID; 7143)
Material examined
Holotype, one female in a permanent, gelation-glycerine glass slide mount (Rot. 54998), paratypes females in permanent slides (Rot. 54999-55004), and 70 females in two glass small bottles preserved in 5% formalin. All the material was collected on 18 January 1989, by A. Adabache-Oritz, C.H. Fernando, and M. Silva-Briano, and is deposited at the Zoological Institute, Russian Academic of Science, St. Ptersburg, Russia. Paratypes deposited at the British Museum (Natural History) London, England, SW7 5BD (1994.4092, 1994.4093, 1994.4093). Paratypes deposited at Royal Belgium Institute of Natural Sciences, Brussels, Belgium, (28101). Additional material: 26 females in formalin deposited at Museo de Zoologica, Escuela Nacional de Estudios Profesionales, Iztacala, UNAM, Mexico, (ENEP)(ROT-48). More material came from a reservoir of Fortaleza, Brazil, collected during August 1989 by C.H. Fernando. (ref. ID; 7143)
Measurements
Total length of lorica 180-192 µm, maximum width 60-80 µm, minimum width 44-64 µm, anterior margin width 60-74 µm. Length of anterior spines median 28-32 µm, intermediate 18-24 µm, lateral 24-28 µm (their tips 12-16 µm). Amictic eggs approximately spherical (52-54 µm), with barely visible dots. (ref. ID; 7143)
Keratella americana nhamundaiensis Koste, 1982 (ref. ID; 2808); Keratella americana nhamundiensis Koste, 1982 (ref. ID; 2806)
Descriptions
This taxon was first discovered in plankton samples collected by G.-O. Brandorff at the mouth of the Rio Nhamunda in July, 1975. It was associated by Koste (Brandorff et al. 1982) to Keratella americana Carlin 1943 species group as a new subspecies. In this species group, however, the hind median facet of the dorsal lorica is open, and in the new taxa this facet is closed, points downwards and ends in a ridge parting to the left, which divided two asymmetrical caudal plaques. Similar although symmetrical, configuration of the caudal facet is found in Keratella procurva (Thorpe 1891), which was not encountered in the study area. K. procurva has been found in the Neotropics in Argentina, Panama and Venezuela. It has a pair of hind spines of which the left is always shortened or lacking. It is possible that, from a morphological point of view, K. nhamunda represents a phylogenetic link between the species groups K. americana and K. procurva. The distinctiveness of K. nhamunda is reinforced by the surface of the dorsal lorica being covered by fine, wave-like, longitudinal striations, a taxonomical character not observed in other Keratella species. (ref. ID; 2808)
Etymology
Its name derives from it first collection site, Rio Nhambunda. (ref. ID; 2808)
Anuraea procurva Thorpe, 1891 (ref. ID; 3031, 3258, 3514); Keratella procurva Berzins, 1953 (ref. ID; 3031); Keratella valga f. valga Edmondson & Hutchinson, 1934 (ref. ID; 3031, 3258); Keratella valga var. procurva Ahlstrom, 1943 (ref. ID; 3031, 3258) or (Thorpe, 1891) (ref. ID; 3514)
Descriptions
The facets on the lorica are very characteristically; the fourth (from proximal end) median facet, is pentagonal and the more distal facet is divided into two square (almost) facets. (ref. ID; 1847)
Dorsum with three median plaques; posteromedian plaque pentagonal and terminates in short median line which extends up to posterior end of lorica. Posterior spines small and subequal. (ref. ID; 2704)
Lorica roughly dotted. A double row of dots on ventral plate occipitally median. (ref. ID; 3023)
Lorica terminating in two unequal and divergent posterior spines, the right spine being somewhat longer. Anterior dorsal margin with six spines; medians longest and stoutest, curved ventrally; laterals usually a little longer than intermediates. Foundation pattern of the dorsal plate characterized by a series of median plaques. Three median plaques present behind the anterior median area. All median plaques completely enclosed. Postero-median plaque pentagonal, terminating in a short median line. Three pairs of enclosed lateral plaques present. Three pairs of latero-marginal, triangular-shaped plaques also present. (ref. ID; 3180)
Measurements
Lorica length 80; maximum width 52; right posterior spine 20; left posterior spine 12 µm. (ref. ID; 1805)
Lorica length 88; maximum width 68; left posterior spine 18; right posterior spine 20 µm. (ref. ID; 2704)
Total length 170; length of lorica 88; lorica width occipitally 55; lorica width posteriorly 41; lorica maximum width 65; occipital median spines 28; occipital mediolateral spines 19; occipital lateral spines 17; posterior spine, left 40; posterior spine, right 50 µm. (ref. ID; 3023)
The general shape of the lorica is of pyriformed rather than vaseshaped. A fundamental geometric pattern of the lorica is almost the same as procurva Thorpe, 1891, Hauer, 1937, Ahlstrom, 1944 and Berzins, 1953. But, it is distinguishable from procurva s. str. in 1) the outline of the lorica, 2) position, direction and length of the posterior spines, which are rudimentary and barely variable throughout season in contrast to procurva s. str, 3) smaller size and 4) period and locality of occurrence. As regards affinity with robusta Koste et al. 1980 nothing to say, for not a single photograph has hitherto been shown. (ref. ID; 2874)
Type locality
Hateruma Jima site 5. (ref. ID; 2874)
Measurements
Total length (TL) 120; lorica length (without spines) 80; widest part (WP) 55-70; WP/TL 0.69-0.76; width between caudal spines 28-30; occipital median sinus 20-22; occipital intermediate sinus 18; right posterior spine 5-7; left posterior spine 1-2 µm. (ref. ID; 2874)
Short, squat, relatively wide and opaque lorica, ornamented with large pustules. Facetted as in f. typ., although with minor differences in populations from widely separated localities. All lorica spines, particularly short posterior spines, with wide bases. Latter curved backwards in lateral view. Forms without caudal spines or with only a single posterolateral spine rudiment as in K. tropica (Apstein, 1907) not recorded for K. procurva. (ref. ID; 2758)
Comments
Comparative morphological characteristics and measurements of known forms of K. procurva are as follows. In common forms the lorica surface is smooth or weakly granulated. Total lorica length from 155-220; lorica width 67-80; anterior spines 19-20, 17-22 and 30-40; posterior spines 22-24 (left) and 27-64 (right) µm. K. procurva robusta has a strongly studded lorica surface, total length 148-150; lorica width 82-90; anterior spines 15-17, 10-15 and 22-23; posterior spines 14-15 (left) and 20-23 (right) µm. (ref. ID; 2758)
With or without one or two posterior spines. The lorica is wider in the posterior width than in the anterior portion. The length of posterior or spines varies greatly, from nearly as long as the body proper to entirely lacking. Various type of the species found in Japan are as follows: (ref. ID; 2393)
1. typical type - Posterior spines about 1/2 of body length, both spines almost equal in length and in parallel direction each other; both reticulate and punctate patterns well-developed on dorsum.
2. divergens type - Posterior spines long, more than 1/2 of body length, strongly divergent, both reticulate and punctate patterns well-developed on dorsum.
3. frenzeli type - Posterior spines long, more than 1/2 of body length, nearly equal in length, almost parallel, slightly incurved; dorsal patterns not distinct.
4. testudo type - Posterior spines short, divergent slightly.
5. irregularis type - Right posterior spine well-developed, left one rudimentary.
6. Intermediate type between typical and divergens-type.
7. Intermediate type between typical and testudo-type. (ref. ID; 2393)
Six anterior spines, reticulate sculpture forming a median series of undivided hexagons, maximum breadth of lorica slightly greater than the posterior breadth, the latter greater than the anterior breadth, two posterior spines which are usually subequal, or one asymmetrically placed spine, or without spines. (ref. ID; 3513)
f. platei (Jagerskiold): Posterior spines long, strongly divergent basally, bent round so that apical they lie at right angles to the long axis of the body. An exuberant development of f. divergens from the Baltic.
f. divergens (Voigt): Posterior spines long, more than half the length of the lorica (excluding anterior spines), subequal and strongly divergent, reticulate and punctate sculpture well developed. Syn. var. longispina Thiebaud (1911).
f. frenzeli (Eckstein): Posterior spines more than half length of the lorica, Subequal ad somewhat sinuate, but with parallel axes. Sculpture practically absent.
f. quadrata (Muller): Posterior spines about half the length of the lorica, subequal and subparallel, reticulate and punctate sculpture well developed.
f. testudo (Ehrenberg): Posterior spines short, about one-seventh of the length of the lorica, often somewhat divergent. Punctate and reticulate sculpture both present.
f. brevispina (Gosse): Posterior spines short and equal as in testudo, often somewhat divergent. Punctate sculpture absent.
f. valgoides f.n.: Right posterior spine well developed, left absent. Punctate sculpture absent. Syn. valga Fadeev, 1927, p.p. Wesenberg-Lund 1930 etc. nec. Ehrenberg.
f. irregularis (Jakubski): One posterior spine (right in typical figures absent, the other, represented by a minute papilla).
f. curvicornis (Ehrenberg): No posterior spines, punctate sculpture absent.
Of the other forms described and probably referable to this species. Anuraea squamula Ehrenberg is clearly a very reduced curvicornis without reticulate sculpture. However, it is very doubtful that this is really the same as Brachionus squamula Muller, the anterior spines of which suggest a very round reduced form of Notholca striata, in all probability the use of this name is superfluous. A. falculata Ehrenberg and A. quadridentata Ehrenberg are doubtful forms which probably need not be recognized. (ref. ID; 3513)
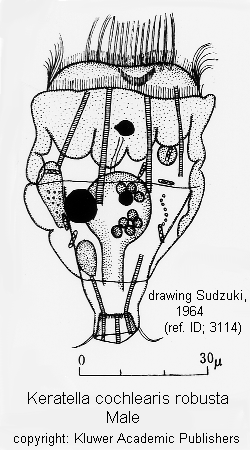
Male: Male present. (ref. ID; 3064, 3065)
Quote from ref. ID; 3064
Measurements
Total length 250-341; the projection of the posterior spines 75-123 µm. The largest individuals are found in early summer. Through June-August a lesser or larger part of the population, up to 1/3, appears as Keratella "canadensis" Berzins. Despite careful measurements it was not possible to separate this form from the species. Intermediate forms can be found with respect to every single character, shape of body and spines as well as relative proportions, angles, etc. Ahlstrom (1943), in his plate XI, Fig.4. gives a picture of a form of K. quadrata, obviously identical with Berzins's K. canadensis. (ref. ID; 1402)
Length of lorica 140-160; width of lorica 86-90; length of median occipital spines 28-35; length of lateral occipital spines 20-26; length of posterior spines 40-55 µm. (ref. ID; 3275)
The form of lorica recalls quadrata divergens Voigt or still more Keratella canadensis Berzins. The lorica is very much narrower occipitally, and very broad posteriorly. The six occipital spines are long and relatively thin, but the posterior spines are very long and thick. The basic pattern in the middle row on the dorsal plaque is first hexagonal; the second field is almost hexagonal, the reverse border is very short. The last pattern, on the posterior border, is pentagonal. From this last nearly hexagonal pattern extends to the base of the posterior spine a hexagonal, elongated lateral field, ending in a broken margin posteriorly. This is peculiarity that separates this new subspecies from the nominate form. (ref. ID; 3023)
Type locality
Taylor Lake, Victoria, Australia. (ref. ID; 3023)
Measurements
Total length 155; length of lorica 122; lorica width occipitally 67; lorica width posteriorly 103; occipital median spines 38; occipital mediolateral spines 34; occipital lateral spines 34; posterior spine, left 105; posterior spine, right 110 µm. (ref. ID; 3023)
According to Sudzuki (1964, pp.31-33: see ref. ID; 3114) two well known subspecies have hitherto been distinguished in Keratella quadrata, namely quadrata s. str. for the Winter specimens without a postero-median remnant and tropica for the Summer specimens with a postero-median remnant. This means that the one criterion of the subspecific characteristics might depend on whether the last plaque of the lorica has a postero-median remant. That is to say, pentagonal (quadrata s. str) or hexagonal (tropica). On the other hand if we adopt Berzin's diagnosis we will arrive at the different conclusion, for Berzins insisted such new criteria as the shape of the lorica and the features of the spines are good to separate the species. The outline of the lorica of the present specimens is very much narrow at the occipital region and rather very broad at the posterior region especially at the basal region of the spine. And, the spines are always thick and divergent just like Berzins' drawing. Thus, the specimens of Berzins seem to be valid. Now, problem may arise as regards the degree of the importance of the characters at the subspecific level. If the specimens with hexagonal pattern occur as the case of valga group, what should we do? Should we regard such a specimen as forma? In the present situation of the knowledge the present writer would like to consider that Keratella quadrata austalis Berzins could be a good species different from quadrata at the specific level. Perhaps Keratella quadrata dispersa Carlin reported by Thomasson in 1955 may belong to this species. (ref. ID; 2417)
Keratella quadrata var. edmondsoni Ahlstrom, 1943 (ref. ID; 3181) reported author and year? (ref. ID; 3114)
Descriptions
The lateral plaques on some specimens were not complete, and the left posterior spine almost obsolete. (ref. ID; 3181)
Measurements
Length overall 220; width 100 µm. (ref. ID; 3181)
Keratella sancta Russell, 1944 (ref. ID; 680, 1923, 3224 original paper, 3521) or 1945 (ref. ID; 2277)
Descriptions
This species has so far only been found in alkaline (pH 9-10) water. The taxon is mentioned by Russell (1960) in his index of the Rotatoria of New Zealand and outlying islands from 1859-1959. Voigt (1957), Kutikova (1970) and Koste (1978) list only the name K. sancta without any remarks, as a rotifer unknown in Europe and the USSR. K. sancta has a heavier pustulation of the dorsal plate. The most closely related species are Keratella cruciformis and K. eichwaldi, because the marginal and endcarinal facets seem to be similar. However, these are marine taxa with a convex caudal lorica and without posterior spines. (ref. ID; 680)
Lorica tub shaped, moderately compressed dorso-ventrally, and with its greatest width approximately three-quarters of the body length (without spines). Anterior dorsal margin with six stout spines, the medians being the longest, converging, and with the distal ends bent in a ventral direction. Intermediate spines diverging, and less than half the length of the medians. The lateral spines very slightly converging to straight, and the same length as the intermediates. Two posterior spines, short, straight, diverging, and inclined ventrally. The foundation pattern of the dorsal plate is unlike that of any other species of Keratella. There are four enclosed plaques, two median, and two lateral: the anterior median plaque is a quadrilateral, and posterior to this is a hexagon. Immediately behind the hexagon are two lateral plaques divided longitudinally by a median line, the division between the hexagon and the lateral area having the form of the letter Y. The foundation patter of the dorsum is very constant between species: in a few cases the division between the anterior median and hexagonal plaques is indistinct, but in every specimen examined it was present. The dorsal plate is postulated, and in the median frontal and marginal areas is heavier than in the medial region. The ventral plate may, or may not, be pustulated. (ref. ID; 3224)
Type locality
This species has so far only been found in Victoria Lake, Christchurch, New Zealand, a lake approximately eight acres in extent, and supplied form deep artesian wells. (ref. ID; 3224)
Measurements
Lorica length 110-200 µm; lorica width 60-120 µm; lorica height 60-80 µm; posterior spines 5-15 µm; median anterior spines 15-37 µm; subitaneous egg 96-72 µm. (ref. ID; 680)
Length of body 135; max. width 95; anterior spines: median 34, intermediate 19, lateral 18; posterior spines 10 µm. (ref. ID; 2277)
The length of the body and spines is very uniform between individuals, as the following. The maximum and minimum values are given from the measurement of six individuals. Length of lorica (without spines) 130-140; maximum breadth of lorica 90-100; length of spines, median 32-37, intermediate 17-21, laterals 16-20, posterior 8-12; width between posterior spines 75-100 µm. (ref. ID; 3224)
Length of body 120, maximum width 90; length of anterior spines: lateral and intermediate 16, median 24; length of posterior spines 10 µm. (ref. ID; 3521)
This species has six anterior spines, two of which are longer and situated in dorsomedian positions. Four shorter spines of equal size occur in dorsolateral positions. Two short posterior spines are also present laterally. The surface of the lorica is covered by evenly scattered pustules which give it a spiny appearance. The pustules are cone-shaped or cylindrical. Furthermore, they are serrated, i.e. longitudinally striated. Their apical end appears to have been cut off, and in its centre there are small knobs and ridges. On the dorsal side the pustules are relatively low, in comparison with those of the anterior par of the ventral plate, which are very long and narrow. They are most numerous in front of transversal ridge in the middle of the ventral plate. Between the pustules, but also on the facet-forming ridges, very small star-shaped structures are seen on both plates. The whole surface of the lorica, except the spines, is covered by a dense network of spot-like structures. (ref. ID; 2214)
Both the punctate and reticular sculptures are remarkable in the dorsal plate. The last central plate is hexagonal, and from its posterior corners a central line runs posteriorly. Both posterior corners project slightly. The central pair of the frontal processes are long and stout, bending downward remarkably near the tip. This is a characteristic species of dystrophic water. (ref. ID; 3063)
Only a vague pattern on the anterior half of the lorica. (ref. ID; 3275)
Comments
Ehrenberg (1858) described under the name Anuraea serrulata two forms which had two medially divided polygons in the rear, the third median polygon being a heptagon. One of these forms had two short posterior spines and was designated as the typical form, while the other had no such spines and was called a variety, but no varietal name was given to it. Hudson and Gosse (1886) also figure two such forms without attempting to differentiate them. A form with long posterior spines occurred in two of the present collections from Maine. While there are probably intergradations among the three forms, it now seems convenient to distinguish them by varietal names.
K. serrulata (Ehrenberg) - There are two short posterior spines. In the Ehrenberg's figure (1838) they are one-twelfth of the total length, or one-third of the length of the anterior median spines. It would be most convenient to regard this simply as a transition between the spineless and long-spined forms (described below), but Ehrenberg definitely established this as the typical form, and the spineless one as a variety.
K. serrulata var. aspina n. var. - The posterior spines are completely lacking. The posterior margin is convex.
K. serrulata var. ehrenbergi n. var. - The posterior spines are long, being as long as the anterior median spines, or about one-fifth of the tool length. (ref. ID; 2771)
Measurements
Length of lorica (with spines) 145; width of lorica 72; length of median occipital spines 40; length of lateral occipital spines 30 µm. (ref. ID; 3275)
Keratella serrulata f. curvicornis (Rylov) (ref. ID; 1472), var. curvicornis Rylow, 1926 (ref. ID; 2593, 3688)
Synonym
Keratella serrulata var. aspina Edmondson, 1936 (ref. ID; 3688)
Descriptions
The most common variety of this acidophilic species, lacking posterior spines, showing a narrow and truncated posterior margin. The middle anterior dorsal spines are bent ventrally. All spines carry fine sharp spicules, just as the anterior ventral margin. The dorsal plate is pusturated. (ref. ID; 2593)
Measurements
Total length 190-192; width 85-102 µm. (ref. ID; 2593)
Keratella serrulata var. aspina Edmondson, 1936 (ref. ID; 2771 original paper, 2836)
Total length 200; length of anterior median spines 48; length of posterior spine 45; width of body 80 µm. (ref. ID; 2771)
Keratella shieli Koste, 1979 (ref. ID; 2757, 2891 original paper)
Descriptions
Very sturdy lorica of approximately trapezoid outline, with a strongly arched dorsal surface. Caudal plate present, connected in middle to convex ventral plate. 6 apical spines medium-sized and strong; 2 short but pointed caudal spines present. Strongly granulated dorsal lorica surface divided by cuticular ridges into facets and panels. These are (unpaired) a median, deep, downward-drawn frontal panel, a rectangular anteromedian panel, a normal hexagonal mesomedian panel, and a trapezoid posteromedian panel which forms a basis for the semicircular caudal plate through a more or less median, arched, strongly ridged frame. Paired panels present: a triangular median and 2 longish lateral marginal panels, and only 2 polygonal lateral panels. Short, very sharp caudal spines seated on a button- to cone-shaped base. (ref. ID; 2891)
Comments
The new species is morphologically distinguished from others of the quadrata species group through the rectangular median panel, the trapezoid rear median panel, the fusion of the anterior lateral panels and the caudal plate. (ref. ID; 2891)
Etymology
This species has been named after Mr R.J. Shiel, Zoology Department, University of Adelaide, in gratitude for ready access to plankton samples from South Australia. (ref. ID; 2891)
Keratella cochlearis var. after Sudzuki (1964) (p.112, plate 18, Figure 9) (ref. ID; 1898); Keratella cochlearis f. tecta after Turner (1986) (p.6, Figure 2a) (ref. ID; 1898)
Diagnosis
By the total absence of any pattern on the lorica, and by the strongly reduced antero-lateral spines, the taxon can hardly be mistaken for any congener. It has been confused with Keratella cochlearis f. tecta. In the latter, the antero-lateral spines are well-developed and rod-shaped, whereas these spines are reduced to small, rounded projections in Keratella sinensis. Additionally, the lorica of K. sinensis appears more elongate than that of K. cochelaris f. tecta. (ref. ID; 1898)
Descriptions
Female (parthenogenetic and mictic female morphologically indistinguishable): Lorica relatively soft, almost twice as long as wide, nearly parallel-sided or weakly constricted in the anterior half. Dorsal plate transversally curved, smooth. Median pair of anterior spines well-developed, nearly straight in lateral view, intermediate pair shorter than median spines, occasionally reduced. Lateral pair reduced to small rounded or triangular projections. No caudal spines developed, even in the presence of predatory Asplanchna species. Apertures to the lateral antennas approximately half-way along the lorica, ridge around the aperture consisting of fused pustules. Ventral plate weakly ornamented anteriorly, otherwise smooth, anterior margin bilobate. Trophi malleate, nearly symmetrical. Unci-teeth six, fused; anterior teeth strongest, gradually declining to posteriorly. Manubria with posterior lamellae prolonged along the shaft. Rami with well-developed inner projections consisting of fused, minute teeth, small allulae present. Fulcrum rod-shaped proximally, dilating distally to a basal plate. (ref. ID; 1898)
Male: Small, illoricate, with large foot and penis. Digestive tract rudimentary, no trophi. (ref. ID; 1898)
Keratella valga slacki Berzins, 1963 (ref. ID; 1834, 2757)
Descriptions
K. slacki shows considerable variations with respect to both lorica length and posterior spine development. Size of the substaneous eggs is relatively constant, so that females of 520 µm and also females of 240 µm carry eggs of 80-84x60-64 µm on the posterior border of the dorsal lorica. (ref. ID; 1834)
Measurements
Total length 205-305 µm; lorica length 120-150 µm; maximum width 60-90 µm; anterior median spine length 40-60 µm; left posterior spine length 40-50 µm; right posterior spine length 60-110 µm. (ref. ID; 1709)
This North American endemity is not particularly rare and is found regularly in large oligotrophic lakes. Occasionally a misformed specimen was seen where one or both lateral 'antlers' pointed ventrally. (ref. ID; 2593)
The dorsal pattern in the lorica was clearly visible. Posterior spine very long. (ref. ID; 2715)
Ahlstrom (1943) has indicated the varying degree of divergence of the antero-lateral spines in Keratella taurocephala. This phenomenon was also noted in the present study and the degree of divergence varied in specimens from different localities and even among specimens from the same sample. (ref. ID; 3086)
Comments
Keratella taurocephala is recorded for the first time from outside North America, and is a new record for Sri Lanka. (ref. ID; 2715)
Measurements
Total length 285; length of body 109, width 83; anterior lateral spine 54; anterior intermediate spine 16; anterior median spine 38; posterior spine 125 µm. (ref. ID; 2715)
The lorica of this species is stout, broad, faintly dotted, the frontal and caudal spine blunt, powerful, and about the same length. It is distinguished easily from K. quadrata by an extra pair of lateral fields, which form a cross-pattern with the last two median fields. (ref. ID; 2597)
Lorica elliptical without posterior spines. Anterior median spines longer. Dorsal plate of lorica with 3 completely formed fields along median axis; first field square like, forth incomplete. Posterior end of lorica with row of marginal fields. (ref. ID; 1806)
Anuraea aculeata var. tropica Tschugunov, 1921 (ref. ID; 3031) or Tschungunoff, 1921 (ref. ID; 3688); Anuraea valga f. tropica Apstein, 1907 (ref. ID; 3031, 3688), var. tropica Apstein, 1907 (ref. ID; 3514); Keratella quadrata Spandl, 1926 (ref. ID; 3031, 3688); Keratella quadrata valga f. asymmetrica Ueno, 1938 (ref. ID; 3031, 3688); Keratella valga f. asymmetrica Hauer, 1937-1938 (ref. ID; 3688); Keratella valga f. brehmi Ahlstrom, 1943 (ref. ID; 3031); Keratella valga f. tropica Edmondson & Hutchinson, 1934 (ref. ID; 3031), Hauer, 1937-1938 (ref. ID; 3688) or Ueno, 1938 (ref. ID; 3688)
Descriptions
Keratella tropica is a loricate rotifer widely distributed in tropical waters and extends very often into subtropical areas as a summer form. A typical form of the species bears two posterior spines on the lorica with the left spine shorter than the exuberant right one. The three morphological forms described.
Form I Keratella tropica f. reducta: This form was present in January and December 1980 and in December 1981. (ref. ID; 663)
Form II Keratella tropica f. asymmetrica: This form was observed in January 1980 and January and February 1981. (ref. ID; 663)
Form III Keratella tropica f. heterospina: This form was present in February and March 1980 as well as in February and march 1981. Few specimens wer also obseved in April 1981. (ref. ID; 663)
The above account indicates clearly that during its morphological seasonal change K. tropica increased in body size and length of the spines through winter to spring. Besides, there was an increase in the length ratio between left and right postero-lateral spines. (ref. ID; 663)
Pejler (1962) suggested that these variations in Keratella tropica may by geographical, discontinuous or non-adaptive. Later (1980), Pejler considered some environmental factors as temperature, food availability, a dilution factor and interspecific biotic relationships to cause form variations in rotifers. (ref. ID; 670)
This species show morphological (cyclomorphic) variation. (ref. ID; 1803)
The posterior spines have varying lengths. However, the left is always shorter than the right. A diminutive form of this species with small lorica and small stumpy spines was found in a sample from a river (Batuluoya). This may be a special adaptation for running waters (Hynes 1970). (ref. ID; 2715)
Lorica compressed dorsoventrally. The anterior dorsal margin with six spines, medians being the longest, stoutest and curved ventralward. Lateral usually little longer than intermediate. Lorica with two unequal posterior spines. Right spine being always longer than the left. The left posterior spine is being absent in some specimens. The dorsal surface of lorica consists of three median polygones and a small four sides structure and three pairs of marginals. (ref. ID; 2867)
Comments
Kratzschmar (1908) confirming the observations of Lauterborn (1900, 1904) reported that long spined forms hatched out both from resting eggs and in parthenogenetic individuals. In his studies the spine tended to shorten and was completely lost successively influenced by internal factors. Klausener (1908) and Hartmann (1918) maintained that external factors, too, might have modified the extent of variation. Green (1960) noted that length of left postero-lateral spine first increased and then decreased to a very short length from February to April. Arora (1966) oberved four forms of K. tropica. In his study the variation involved reduction and ultimate disappearance of the left posterior spine. The right spine also started reducing until a form comparable to reducta was produced. Finally, this spine also disappeared. Ultimately, the cycle started from heterospinate form and reached f. aspina through f. reducta. However, this sequence was reversed in K. valga. Green (1980) investigated asymmetry and form variation in K. tropica from some tropical lakes and found that there was a general tendency for the length of the right spine to incease with the length of lorica. In the present study three morphological forms of K. tropica were distinguished: f. reducta, f. asymmetrica, and f. heterospina. The cycle started, however, from f. reducta characterized by presence of a small right postero-lateral spine and absence of a left postero-lateral spine or presence of a very small one. Many authors considered this phenomenon to be the effect of an increased temperature in summer (Lauterborn 1900, 1904; Wesenberg-Lund 1900, 1930; Ahlstrom 1943; Gallagher 1957) or spring (Carlin 1943). Baker (1979) also observed increased spine lengths of K. cochlearis and K. earlina with increasing temperature. Buchner et al. (1957) on the other hand found long appendages at low temperatures and short ones or none at higher temperatures. Amren (1964) could not find any apparent connection of seasonal variation in K. quadrata with temperature. In our study, temperature seemed to be one of the factors responsible for form variation in K. tropica as f. reducta appeared in December, f. asymmetrica in January and February and f. heterospina in February, March and April when temperature increased continuously. Most of the forms with long spines described in several studies have been observed so far at low temperatures in temperate climate. Edmondoson (1948) pointed out that calcium might have been an effect on seasonal variation. (ref. ID; 663)
This species was separated from K. valga (Ehrenberg) by Berzins (1955). The distinguishing character, i.e. the hindmost median plaque of the dorsal lorica (the "postero-median remnant" according to the terminology used by Ahlstrom 1943), is clearly discernible in most individuals, although in some specimens from L. Naivasha (on July 18, 1968) it was very small and hardy visible. (ref. ID; 3020)
Measurements
Lorica length 120; maximum width 75; left posterior spine 32; right posterior spine 85 µm. (ref. ID; 2704)
Total length of lorica 240 µm. River form; total length 132; width 63 anterior lateral spines 18; anterior intermediate spines 12; anterior median spines 30; right posterior spine 18; left posterior spine 12 µm. (ref. ID; 2715)
Total length 250; length of lorica 130; maximum width 90; length of antero-median spine 45; length of antero-lateral spine 30; length of right posterior spine 90; length of left posterior spine 30 µm. (ref. ID; 2867)
Keratella tropica f. asymmetrica (Barrois & Daday) (ref. ID; 2619, 2628, 3688) reported author and year? (ref. ID; 663)
Synonym
Anuraea valga f. asymmetrica Barrois & Daday, 1894 (ref. ID; 2628); Keratella valga f. asymmetrica Hauer, 1938 (ref. ID; 2628)
Descriptions
Size increased to 252 µm. The sizes of left and right posterior spines increased to 42 and 98 µm, respectively. (ref. ID; 663)
The typical fully developed form, measuring 280 µm in total length. The size of left and right posterior spines increased 56 µm and 112 µm, respectively. (ref. ID; 663)
This specimens has only one posterior spine. (ref. ID; 2704)
Keratella tropica f. reducta Fadeew, 1927 (ref. ID; 3688) reported author and year? (ref. ID; 663)
Synonym
Keratella quadrata var. tropica f. reducta Fadeew, 1927 (ref. ID; 3688)
Descriptions
Size up to 238 µm. A very small left posterior spine appeared, measuring 28 µm whereas the length of the right posterior spine was 84 µm. (ref. ID; 663)
Keratella valga (Ehrenberg, 1834) (ref. ID; 1345, 1350, 1803, 2277, 2393, 2757, 2816, 2836, 3031, 3114, 3264, 3688) reported year? (ref. ID; 3029, 3065, 3126, 3208, 3484, 3513, 5022, 5031) reported author and year? (ref. ID; 2871, 3064), valga f. valga Ehrenberg (ref. ID; 3523), valga valga (Ehrenberg, 1834) (ref. ID; 1345, 2860, 3073, 3083)
The width of body proper is greater at the anterior end than at the posterior end. The posterior spines are unequal in length; the left is shorter than the right. Sometimes, the left spine is absent, even if the right one is well-developed; both spines are absent in some cases. Both reticulate and punctate patterns on the dorsum are fundamentally similar to those of K. quadrata. Various length of posterior spines have been observed by many investigators for taxonomic purposes. In this respect, Edmondson & Hutchinson (1934) described various varieties and forms of this species. Ahlstrom (1943) who did not distinguish various forms for the present species on the basis of only the length of posterior spines, gave two groups of variants in the feature of typical patterns on the dorsum, and concluded that it was divisible to two definite forma names, f. tropica and f. brehmi. The variants found in Japan belong to Ahlstrom's f. tropica, which may be identical with the following six forms offered by Edmondson & Hutchinson. (ref. ID; 2393)
1. typical type - Posterior spines developed, left spine about 1/4, right spine about 1/3 of body length.
2. asymmetrica type - Right posterior spine about 1/2 of body length, left one very short.
3. tropica type - Right posterior spine about 6/7, left one 2/5 of body length.
4. monstrosa type - Right posterior spine about 3/5 of body length, left one lacking.
5. tropica-asymmetrica type - Right posterior spine long, left one very short.
6. tropica-monstrosa type - Right posterior spine long, left one lacking. (ref. ID; 2393)
Normal specimens with long anterior spines and a long right posterior spine. The left posterior spine absent. (ref. ID; 3126)
Six anterior spines, reticulate sculpture forming a medial series of undivided hexagons, maximum breadth of lorica very distinctly greater than the posterior breadth, the latter much less than the anterior breadth, two posterior spines which are usually unequal, or one asymmetrically placed spine, or without spines. (ref. ID; 3513)
f. valga (Ehrenberg): Posterior spines well developed, rather short, unequal, the right typically one-third and the left one-fourth of the length of the lorica. Puncatate and reticulate sculpture both present.
f. asymmetrica (Barrois & Daday): Right posterior spine well developed, typically about half as long as the lorica, left spine rudimentary. Punctate and reticulate sculpture both present. Syn. f. heterospina Klausener (1908) p.p.
f. monstrosa (Barrois & Daday): Right posterior spine well developed, typically about half as long as the lorica, left spine absent. Reticulate and punctate sculpture well developed. Syn. ?var. dumasi Richard (no figure or indication if this is really a form of valga); var. asymmetrica Daday nec. Barrois & Daday; f. monospina Klausener (1908); f. monospina Fadeev (1927) = tropica-monstrosa.
f. longicornis (Schmarda): Both posterior spine well developed and practically equal, typically about three-fifths as long as the lorica, reticulate and punctate sculpture both well developed, as indicated in Schmarda's typical figure, or the latter reduced (Hartmann 1918, Figure 76).
f. tropica Apstein: Right spine very long, typically about six-sevenths of the length of the lorica, left spine much shorter but well developed, about two-fifths as long as the lorica. Reticulate sculpture developed, punctate often reduced.
f. reducta Fadeev: Right posterior spine alone developed, short, about one-fourth of the length of the lorica (typical figure may be taken as Fadeev 1927, T.2, fig.12), punctate sculpture absent, reticulate very reduced.
f. brehmi Klausener: Both posterior spines absent, reticulate sculpture present (Brehm & Zederbauer 1904, fig.1), punctate apparently absent.
f. aspina Fadeev: Both posterior spines absent, reticulate sculpture almost and punctate entirely absent. (ref. ID; 3513)
Male: Male present. (ref. ID; 3064, 3065)
Quote from ref. ID; 3064
Comments
In true K. valga the cycle (Klausener 1908; Hartmann 1918) appears to involve primarily the addition of spines. The initial phase in the most complete cases is the form called by Klausener K. curvicornis f. brehmi. This form is figured as being somewhat longer than K. quadrata f. curvicornis. In certain irregularities in the line of the posterior margin of this figure there is, moreover, perhaps a hint of the position of the spine bases, demarkating a typical valga posterior margin, though this may be accidental. Fadeev (1927) has described and figured as f. aspina a comparable form which is certainly clearly referable to valga on the shape of the posterior margin alone without having to take into account the other members of the cycle. An essentially similar elongate form was recorded from South Africa by Hutchinson and by Hutchinson, Pickford and Schuurmann (1932) as curvicornis. Klausener distinguished f. brehmi from curvicornis by the fact that the antero-median spines do not diverge in the former. This character is probably too variable and in general does not separate valga from quadrata. It must be admitted that the two species must often be hard to separate in their most reduced forms. In Hartmann's studies of this species the most exuberant form was one in which the two posterior spines are subequal. This form seems to be the one figured by Schmarda (1850, Plate IV, fig.III) as A. longicornis but it has doubtless been regarded by other authors as aculeata (i.e. quadrata s.s.). Apstein has described another exuberant form from Ceylon as tropica. This form is characterised by the very long and unequal posterior spines, the left being about two-fifths the length of the lorica without the anterior spines, the right about six-sevenths of this length. This form, in spite of Hartmann's statement that it agrees well with his form of May, 1915 (f. valga), differs from his figure of the latter in which the right spine is hardly more than one-half the length of the lorica. It is clear from the work of Tschugunoff (1922) and Fadeev (1927) that forms essentially similar to tropica are common in South Russia, and Skorikov as long ago as 1896 figured (T.VIII, fig.29) such a form from near Kharkov. Jakubski (1915) has figured a monospinous tropica form from Poland and in South Africa, Hutchinson (1930) and Hutchinson, Pickford and Schuurmann (1932) found tropica to be the only common form of valga. In all probability the most developed form of valga varies form place to place, and if f. valga and f. longicornis represent the highest development in some European localities, tropica probably does the same in Ceylon, S. Russia, South Africa and Kashmir. The names proposed for forms of valga in which the left spine is very reduced or absent may be used in conjunction with the name tropica in localities where the right spine is very elongate, and where these reductions occur, e.g., K. valga f. tropica-asymmetrica and K. valga f. tropica-monstrosa. (ref. ID; 3513)
Total length 164-200; dorsal lorica length (without spines) 100-110; width at the basis of occipital lateral spines 56-60; widest part 64-70; height at the base of frontal spines 20-24; highest part 51-53; anterior median sinus 20-38; anterior lateral spines 19-23; pectoral median sinus 15-20 wide, 8 deep; caudal projection or protuberance 0-40 (left), 48-80 (right); fertilized dormant egg 59-60x41-50 µm. (ref. ID; 3083)
This form belongs clearly to valga-tropica group, but, demarked by possessing extraordinarily long posterior spines, especially the right one is longer enough to exceed the body length. No variation has been observed in the length of posterior spines. This is true for all the sites surveyed end in winter at site 4. It is worth mentioning that 1) only from site 1 & 2 on 28 Feb. and 18 Dec., the usual type of tropica were encountered. But, strangely enough they all belonged to f. monstrosa Barrois & Daday, 1894, 2) Ueno (1938) could not observe f. diatojimensis from Kita-Daito Jima, located just on the north of this coral island. (ref. ID; 2874)
Type locality
Minami-Daito Jima site 4. (ref. ID; 2874)
Measurements
Total length (TL) 280-300; lorica length (without spines) 120; widest part (WP) 80-82; WP/TL 0.67; right posterior spine 120-140; left posterior spine 60-65 µm. (ref. ID; 2874)
The form of the lorica is more like that of Keratella tropica (Apstein) than of valga (Ehrenberg), and is very elongated and narrow. But this animal lacks the post-median remnant pattern. There is, however, a rounded ridge towards the posterior margin, extending from the ridge to the base of the posterior spines. Another difference is to be found in the elongated ridge of the median occipital spines. There are two very abrupt margins between extending forward. The occipital median and posterior spines are long. (ref. ID; 3023)
Type locality
Taylor Lake, Victoria, Australia. (ref. ID; 3023)
Measurements
Total length 456; length of lorica 192; lorica maximum width 117; lorica width occipitally 78; lorica width posteriorly 75; occipital median spines 77; occipital mediolateral spines 34; occipital lateral spines 27; posterior spine, left 83; posterior spine, right 190 µm. (ref. ID; 3023)
Keratella valga var. procurva (Thorpe, 1891) (ref. ID; 1345, 2278)
The majority of the specimens had the left spine obsolete or weakly developed. The dorsum pattern was distinct and normal for the species. (ref. ID; 2278)
Measurements
Overall length 200; width 90; anterior spines: AM 37, AI 20, AL 25; posterior spines: left 0-20, right 40-50 µm. (ref. ID; 2278)