The World of Protozoa, Rotifera, Nematoda and Oligochaeta
Hexarthra
Hexarthra Schmarda, 1854
Class Rotatoria: Order Monimotrochida: Family Hexarthridae (ref. ID; 7097)
See Pedalia (This genus was formerly known as Pedalia Barrois (originally Pedalion Hudson), but the priority of Hexarthra is recognized (Neal 1951; Hemming 1955) (ref. ID; 1923)
Synonym Pedalia Barrois, 1878 (ref. ID; 3514, 3688, 4596, 5932); Pedalion Hudson, 1871 (ref. ID; 3514, 3688, 4596, 5932)
ref. ID; 830
The identification of the various species was done preserved material only and was based on the trophi of the mastax. Hexarthra is characterized by a mastax with malleoramate trophi and the number of teeth on the uncus is used in taxonomy. According to Ruttner-Kolisko (1974) the various morphological characteristics (e.g. size, relation of the ventral arm/body length, number of bristles, number of teeth on the uncus) by which the species of this genus are distinguished show considerable variation in different environmental conditions; there seems to be a tendency towards an increase in the number of teeth on the uncus as salinity increases, and the ratio length ventral arm/body length decreases in warmer climates. Nevertheless, according to Koste (1978), the four species can be identified on the basis of the number of teeth and display clear ecological differences. (ref. ID; 830)
ref. ID; 1663
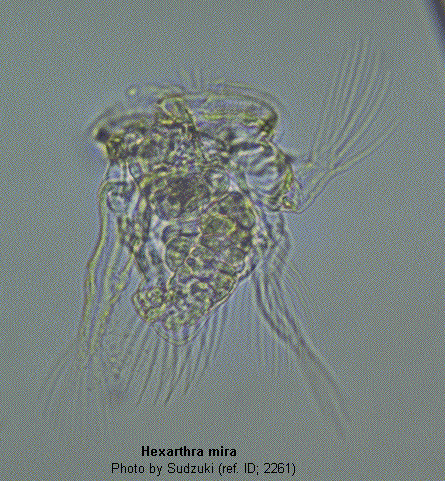
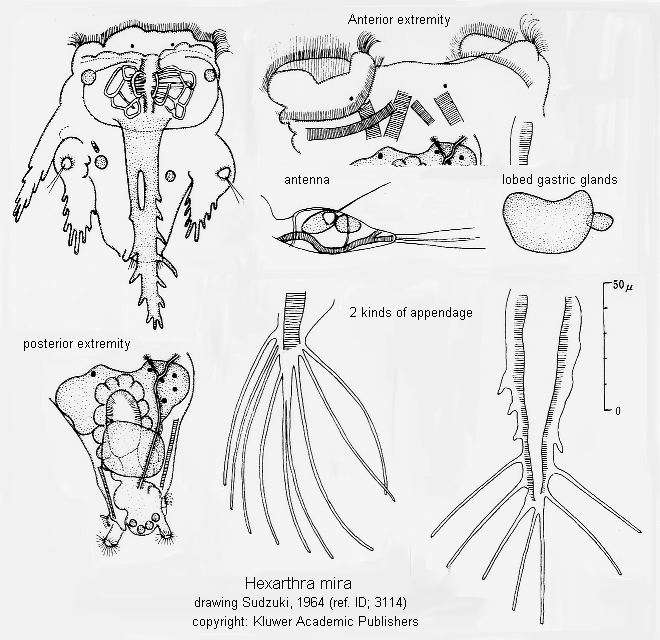
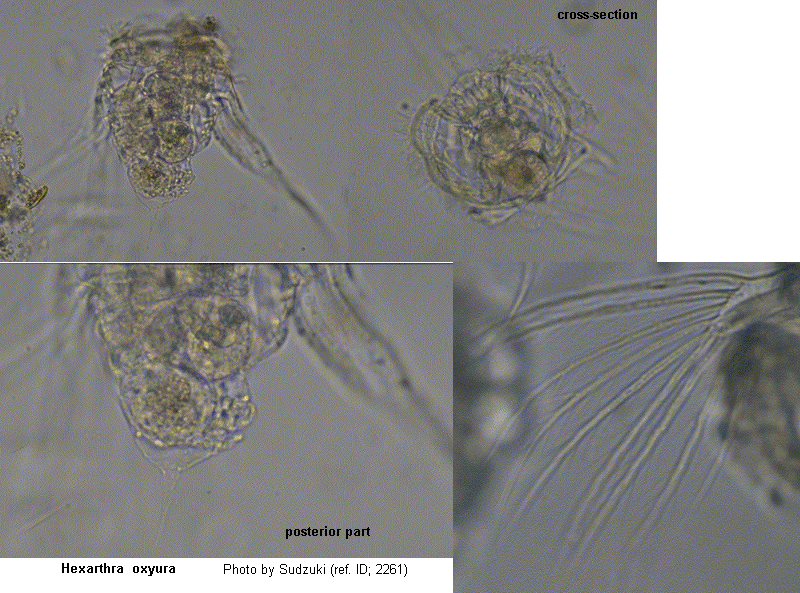
With six stout, muscular, setose appendages. Body conical. With a double ciliary wreath and a ciliated groove between. The "jumping rotifer". Usually in small alkaline lakes and ponds during summer. (ref. ID; 1663)
ref. ID; 3087
The object of the present application is to invite the International Commission on Zoological Nomenclature to use its plenary powers to stabilise the name for the genus of the Class Rotifera formerly known as Pedalion Hudson, 1871, or as Pedalia Barrois, 1878, by designating a recognisable species to be the type species of the genus Hexarthra Schmarda, 1854. The facts of this rather complicated case are set out in the following paragraphs.
1. The generic name Pedalion Hudson, 1871 (Mon.microscop.J. 6: 121, pl.94) was established for a monotypical genus for a new species then made Pedalion mira. The name Pedalion Hudson, 1871, is invalid, because is a junior homonym both of Pedalion Dillwyn, 1817 (Descr.Cat.Shells: 282) and of Pedalion Swainson, 1838 (Nat.Hist.Fishers 1: 199). It will be convenient if, before considering whether there is any subjectively identical nominal genus, the name of which should replace the invalid name Pedalion Hudson, 1871, we examine briefly the subsequent history of Hudson's name Pedalion.
2. In 1877 Barrois gave a paper at the Sixth Session of the "Association francaise pour l'Advancement des Sciences" held at Le Havre, entitled: "Sur l'anatomie et le developpement du Pedalia mira". This paper was published in 1878. Although, as noted above, the generic name was written in the form Pedalia in the title of Barrois' paper, that name appeared in the form Pedalion at ever point at which it appeared in the body of that paper. Rousselet, in 1914 (Association francaise pour l'Advancement des Sciences, 43e Session, Le Havre, C.R.: 535-536) drew attention to this inconsistency and raised the question whether it was due to an editorial error or whether it a should be explained on the ground that Barrois was aware of the fact that the name Pedalion Hudson, 1871, was invalid and tried to get over this difficulty by substituting the variation form Pedalia in the title of his paper. It must be noted however that an abstract of this paper of Barrois' was published in the issue of the Revue scientifique of 29th September, 1877 and that in this paper only the spelling Pedalion occurs. In this abstract no title was given for this paper and it possible that it is for this reason that the spelling Pedalia did not appear on this occasion, for it was only in the title of Barrois' paper that the spelling Pedalia was used. It may be noted incidentally at this point that the species dealt with by Barrois in the foregoing paper is not only not (as he supposed) the species which in 1871 Hudson had named Pedalion mira; it is not even referable to the same genus; it is actually a marine species of the genus Synchaeta Ehrenberg, [1832] (Abh.Preuss, Akad.Wiss, 1831: 135).
3. Since 1913 the genus named Pedalion by Hudson has been called both by that name and by the reputed name Pedalia Barrois, 1878, the latter having even made its way into Neave's Nomenclator zoologicus (3: 632), where it is quoted as a substitute name for Pedalion Hudson, 1871, the reference being given as "Pedalia Barrois, 1878, C.R.Ass.Franc. 6 (1877 Le Havre), 661".
4. I am bound to say that, in my view, it is impossible to accept, as a valid substitute name, a name (such as Pedalia) which appears only in the title of a paper, the correct spelling (in this case, Pedalion) occurring at every point in the body of the paper itself, where, if Barrois had intended to emend Hudson's name Pedalion in this way, he would certainly have used the spelling "Pedalia". It is clear, however, that no progress can be made in the stabilisation of the name of this important genus until this preliminary question has been disposed of. I accordingly ask the International Commission on Zoological Nomenclature to give a ruling that the use of the word Pedalia in the title of Barrois' paper does not constitute the publication of a generic name consisting of this word, the spelling "Pedalia" being due, it must be concluded, to some inadvertence on the part of the editor of Barrois' paper or on that of printer.
5. It is necessary now to consider whether there is any nominal genus possessing a valid name, the type species of which can be regarded as belonging to the genus Pedalion (or Pedalia) as hitherto understood. An examination of the literature shows that there is such a nominal genus, which does, or may, satisfy this condition. This is the genus Hexarthra Schmarda, 1854 (Denskr.Acad.Wiss.Wien. (Math.-natur. K1.) 7 (no.2): 15). This genus in monotypical, its sole species being Hexarthra polyptera Schmarda 1854, a then newly described (: 15) nominal species. This species is therefore type species of the genus Hexarthra Schmarda by monotypy.
6. Our next step must be to consider the claims of the nominal species Hexarthra polypteraSchmarda to regarded as representing a species of Pedalion (or Pedalia). The specimen upon which Schmarda based his description and figures of this nominal species, which was hardly visible to the naked eye, was discovered by him in a pool at El Kab in Egypt. At first Schmarda mistook this animal for a crustacean larva, but the "Raderorgan" and jaws showed that it was a Rotifer. The jaws had seven teeth on each side. Schmarda's type specimen is missing, and the species which he described has not been reported by any subsequent worker.
7. The drawing up his description and preparing his figures of this species, Schmarda was severely handicapped by reason of the fact that he was working under field conditions and had no opportunity for studying the animal at leisure. This stand is taken also by Daday, 1903 (Mikroskopsche Susswassertiere aus Kleinasien.Wien.Sitzungsber. (Math.-Natur. Kl.) 112 (Abth. 1): 139-168) When after the publication of the name Pedalion, the view was advanced that the genus so named as indistinguishable taxonomically from the genus Hexarthra Schmarda, Hudson objected on the ground that Schmarda had described the six plumose appendages as originating from the ventral side, as in nauplius larva, while in his genus Pedalion these appendages were arranged around the animal. Hudson admitted, however, the difficulty of studying the arrangement of the hairs on the appendages of Pedalion species, a difficulty which may be illustrated by the mistakes which he himself made in some of his own figures and descriptions.
8. Daday (1886, Morph. Physiolog. Beitrage zur Kenntniss der Hexarthra polyptera Schmarda. Termeszetraize Fuzetek 10: 214-249, pls.VIII, IX) believed that, in representing the appendages of Hexarthra polyptera as arising from the ventral side of the animal, Schmarda had been influence by the apparent similarity between members of the genus Pedalion and the nauplius larva of Crustacea, a resemblance which, though apparent at first glance, is found on closer examination to be entirely superficial in character.
9. Levander (1894, Beitrage zur Kenntniss der Pedalion Arten mit einer Tafel.Soc.Faun, Flor.fenn. 11: 1-34, figs.1, 2, 3) considered that, if one were to identify Pedalion with Hexarthra, it would be necessary to attribute to Schmarda a lack of observation which, in his view, there was no ground for assuming. Levander quoted the opinion of Plate-that, if Schmarda could show (as he did) the arrangement of the hairs on the appendages (which in Pedalion are found only on the ventral appendage) and the number of teeth in the jaws, he could not possibly have made the mistake of representing appendages as arising from the ventral side of the animal instead of being arranged in a circle around it. Plate concluded that, in view of these facts, the presence of hooks on appendages other than the ventral appendage, and the origin of the appendages from the ventral side of the animal, it was impossible to synonymise Hexarthra with Pedalion. As regards Plate's comment on these lateral hooks on the appendages, it is, however, worth nothing that Daday recorded the presence of similar hooks on the dorsal appendage in his figure of his Pedalion mucronatum Daday, 1909 (Trav.Soc.Nat.Petersb. 39: 9-38, pl.1, figs.2-6), a nominal species which has been identified with Pedalion oxyure Sternov, 1903 (Turkestanskago Otd, imp.russ.georgr.Obsch. 4 (Pt.3): 9, pl.1, figs.2,3), in the figures of which no lateral hooks are shown on the dorsal appendage. This difference between the illustrations given by Sernov and Daday, to which attention has not so far been drawn, has not prevented authors from accepting Pedalion mucronatum Daday and Pedalion oxyure Sernov as being no more than different names for a single species. I have never noted lateral hooks on any but the ventral appendage. Daday's (1909) representation of hooks on the dorsal appendage may be due to incorrect interpretation.
10. Thus against the view that Hexarthra polyptera Schmarda is a species of the genus that Hudson named Pedalion is the observation by Schmarda that the appendages arise from the ventral surface and that there are lateral hooks on appendages other than the ventral appendage. As already noted however Daday himself showed incorrectly the presence of a such hooks in his P. mucronatum (= P. oxyura Sernov). It must be noted also that at times the fold of the integument on the appendages can give somewhat laddered appearance which under low magnification may be mistaken for hooks. The presence of jaws in H. polyptera speaks for its being a rotifer. In view of the absence of posterior digitiform appendages posteriorly (such as are found in Pedalion mira Hudson), Schmarda's species might well belong to the fennicum-group of the genus Pedalion. The arrangement of the hairs on the appendages is very similar to that found in a species of Pedalion which occurs in the Saskatchewan lakes on Canada. The principal objection to the acceptance of Schmarda's species as belonging to the genus Pedalion is therefore his statement that the appendages come from the ventral side of the animal. As regards this, it must be noted, however, that a similar appearance can be obtained by placing a specimen of an undoubted species of Pedalion under a cover slip, the weight of which has flattened the animal (Schmarda's figure seems to have been drawn from a flattened specimen); unless one constantly alters the focus while drawing the animal as a whole or one of its appendages in order to obtain the correct relation, it is very easy to arrive at the same conclusion as that reached by Schmarda. An inaccuracy of this kind on the part of Schmarda would not be surprising if we recall the difficulties with which he was faced, by reason of having to examine his material in the field instead of in a laboratory.
11. The next point which it is important to note is that Schmarda found his species Hexarthra polyptera in abundance in the waters which he examined. This fact suggests that that species, whatever it was, would have been found again by subsequent workers. The species Pedalia fennicum Levander, 1892. (Zool.Anz. 15: 403) has, in fact, been reported from North Africa, where it appears to be rather widely distributed. Recent observations show that this species shows considerable variation in the arrangement of the hairs and some variation in the tooth formula - a fact which may account, in part, for some of the special features shown in Schmarda's figures. Further, Bryce in his paper on the Rotifera of Devil and Stump Lakes (1924, J.Quekett microscop.Club 15: 81-108) mentioned having examined a slide of Pedalia from El Kab (the type locality of Schmarda's species) that had been prepared by Rousselet. I have myself examined a slide of a specimen from this locality that was bought from Rousselet by the United States National Museum. The specimen mounted on this slide is a Pedalion.
12. The conclusion which I draw from an examination of Schmarda's (admittedly poor) description and figure is that the species which he described as Hexarthra polyptera belongs to Hudson's genus Pedalion. This conclusion becomes a virtual certainty, now that we have an actual specimen of a Pedalion from the very pond in which Schmarda took his polyptera but in spite of the abundance of that species noted by Schmarda no specimen of a species belonging to a separate genus recognisable from Schmarda's description as Hexarthra is known from that very restricted locality. Schmarda's description of the arrangement of the hairs on the ventral appendages, the number of hooks on the ventral appendage that he noted, as also the number of teeth (seven) in each uncus, all place the animal described by Schmarda in the fennicum-group of the genus Pedalion.
13. In the light of the foregoing consideration, I reach the conclusion that the nominal genera Hexarthra Schmarda, 1854, and Pedalion Hudson, 1871, must on taxonomic grounds, be regarded as subjectively identical with one another. Accordingly, the name which under the Code should be applied to this genus is Hexarthra Schmarda and, as it is the older of the two names, would still be the correct name for this genus, even if Pedalion Hudson, 1871, were not an invalid homonym. In view of the considerable discussion that has taken place in regard to this question and also of the importance and interest of this genus, I thick that it is important that this subject should now be closed by the name Hexarthra Schmarda being placed on the Official List of Generic Names in Zoology and the invalid name Pedalion Hudson, 1871, on the Official Index of Rejected and Invalid Generic Names in Zoology, together with the reputed, but non-existent, name Pedalia Barrois, 1878. It would be useful also if all other invalid uses of the name Pedalion subsequent to the name Pedalion Dillwyn, 1817 (Class Pisces) were at the same time to be relegated to the Official Index.
14. At this point we encounter a difficulty of quite a different order. The nominal species Hexarthra polyptera Schmarda, though incorrectly and inadequately described, must be regarded as belonging to the highly variable group represented by Pedalion fennicum Levander, 1892. Accordingly, under the Code the trivial name polyptera Schmarda, 1854, being much older than fennicum Levander, would replace the latter name. The trivial name fennicum Levander is however so deeply entrenched in the literature that its replacement by the hitherto contentious name polyptera Schmarda would certainly give rise to confusion and would be open to strong objection. In order to prevent this confusion, I accordingly ask the International Commission on Zoological Nomenclature to use its plenary powers to suppress the trivial name polyptera Schmarda, thereby validating the later name fennicum Levander for the same species. As part of this proposal, I ask the International Commission to use the same powers to designate Pedalion fennicum Levander, 1892, to be the type species Hexarthra Schmarda, in place of the older nominal species Hexarthra polyptera Schmarda, the name of which it is now proposed should be suppressed. The trivial name fennicum Levander, 1892 as published in the binomial combination Pedalion fennicum should then be placed upon the Official List of Specific Trivial Names in Zoology. Incidentally, it would be helpful if at the same time the International Commission were to place on this Official List the trivial names, each an available name and the oldest name for the species concerned, of two other well-known species of this genus. These trivial names are: (1) the trivial name mira Hudson, 1871 (Mon.microscop.J. 6: 121) (as published in the binominal combination Pedalion mira); (2) the trivial name intermedia Wiszniewski, 1929 (Bull.Acad.pol.Sci.Lettr. (Cl Sci.math.Nat.) (B) 1929 (2): 137) (as published in the binominal combination Pedalia intermedia).
15. The specific proposals which I now therefore lay before the International Commission on Zoological Nomenclature are that it should:-
(1) use its plenary powers:-
(a) to suppress, for the purposes of the Law of Priority but not for those of the Law of Homonymy the trivial name polyptera Schmarda, 1854 (as published in the binominal combination Hexarthra polyptera);
(b) to set aside the indication, by monotypy, of Hexarthra polyptera Schmarda, 1854, as the type species of the genus Hexarthra Schmarda, 1854, and having done so, to designated Pedalion fennicum Levander, 1892, to be the type species of that genus;
(2) place the generic name Hexarthra Schmarda, 1854 (type species, by designation, as proposed in (1)(b) above, under the plenary powers: Pedalion fennicum Levander, 1892) (gender of generic name: feminine) on the Official List of Generic Names in Zoology;
(3) place the under-mentioned generic names or reputed generic name, on the Official Index of Rejected and Invalid Generic Names in Zoology:-
(a) Pedalia Barrois, 1878 (a reputed but non-existent name);
(b) Pedalion Swainson, 1838 (a junior homonym of Pedalion Dillwyn, 1817);
(c) Pedalion Hudson, 1871 (a junior homonym of Pedalion Dillwyn, 1817);
(d) Pedalion Buckton, 1903 (Monogr.Membr. 6: 251) (a junior homonym of Pedalion Dillwyn, 1817);
(4) place the under-mentioned trivial names on the Official List of Specific Trivial Names in Zoology:-
(a) fennicum Levander, 1892 (as published in the binominal combination Pedalion fennicum) (trivial name of species proposed, under (1)(b) above, to be designated as the type species of Hexarthra Schmarda, 1854);
(b) intermedia Wisniewski, 1929 (as published in the binominal combination Pedalia intermedia);
(c) mira Hudson, 1871 (as published in the binominal combination Pedalion mira);
(5) place the trivial name polyptera Schmarda, 1854 (as published in the binominal combination Hexarthra polyptera) (a name proposed, under (1)(a) above, to be suppressed under the plenary powers) on the Official Index of Rejected and Invalid Specific Trivial Names in Zoology. (ref. ID; 3087)
ref. ID; 4596
With six, stout, setose appendages. Body conical. Corona with a double ciliary wreath, and a ciliated groove between. (ref. ID; 4596)
Very like H. oxyuris habitually with the same type of corona. But trophy with only 6/6 teeth. Abdominal prolongations varies, can be short or very long and thin. (ref. ID; 4606)
Type locality
Found common rotifer in Lake Barracoota plankton. (ref. ID; 4606)
Measurements
Total length 135; body length 95; Posterior prolongation between 20 and 40 µm. (ref. ID; 4606)
Ruttner-Kolisko (1974) has divided the genus into two main groups of species: the fennica-jenkinae group and the mira-intermedia group and suggested that H. bulgarica occupies an intermediate position between them. However, Koste (1978) prefers to include the species in the "formenkreis" propinquamollis-bulgarica. Dumont et al. (1978) suggested that H. bulgarica can be divided into three subspecies: H. b. bulgarica, H. b. nepalensis and H. b. canadensis. Although these authors note that the only morphological feature common to all these subspecies is the presence of a pair of spines on the dorsolateral arm, certain ecological characteristics help to define the species. (ref. ID; 1449)
H. b. canadensis (ref. ID; 1449)
H. b. nepalensis (ref. ID; 1449)
Descriptions
(1) absence of ventral appendages, (2) absence of lower lip, and (3) presence of spines on the dorsolateral arms. (ref. ID; 1449)
Body conical, no caudal appendages, length 240-270 µm, and no ventro-anterior lip. The ventral arm is longer than the boby. Its spine formula is 5/5 or 5/4 and the number of terminal filaments is 10 to 13, more often 13. They are implanted on five stems. The dorsal arm has six to eight filaments, usually seven. The ventrolateral arm has seven or eight filaments, usually eight. The dorsolateral arm has six or seven filaments, usually seven, plus two basal spines. The unci have a tooth formula 7/7, 7/8, or 8/8. The teeth are not differentiated into larger and smaller ones, but gradually disminish in size dorsally. (ref. ID; 5932)
Remarks
Subspecies canadensis differs from bulgarica and nepalensis in the greater number of teeth on the unci, the larger number of spines on the ventral arm, and the tendency to have at least one additional filament on every arm. In addition, it lacks the ventro-anterior lip present in nepalensis. Hexarthra bulgarica canadensis could be mistaken for H. fennica on the basis of the number of teeth on the unci, although H. fennica has no spines on the dorsolateral arm. the latter character appears to be diagnostic for the bulgarica group. The number of teeth on the unci and the presence or absence of a ventro-anterior lip are characters which vary at the subspecific level. This point of view is substantiated by the habitat selection of H. bulgarica, which is a typical high-mountain, freshwater species. In fact, it is the only truly high-mountain rotifer known. It was described from the Rila Mountains, Bulgaria (Wiszniewski 1933), but is found also in the Himalayas and, according to Ruttner-Kolisko (1972, noting personal communication with H. Loffler) also in the Andes. The status of the Andes form is unknown, but on geographical grounds it could be the same as, or similar to, H. bulgarica canadensis. (ref. ID; 5932)
Type materials
Holotype: Undissected female mounted in glycerine on a slide, deposited at the KBIN/IRSNB. (ref. ID; 5932)
Paratypes: Ten whole animals mounted individually in glycerine on slides, five mastax preparations, and about 50 specimens preserved in a formaldehyde-glycerine solution. Half the paratype material is in the Laboratorium voor Dierkunde, Rijksuniversiteit Gent, Belgium, and half in the Canadian Wildlife Service laboratory, Department of Biology, University of Calgary, Canada. (ref. ID; 5932)
Material examined
About 80 specimens, all parthenogenetic females from Grouse Lake, Alberta, Canada, 11 July 1968, and 27 July 1968, R.S. Anderson leg. (ref. ID; 5932)
A full description is given by Daems and Dumont (1974) under H. bulgarica (Wiszniewski 1933). The main distinctive character is the ventro-anterior lip which is very strongly developed in H. bulgarica nepalensis, but is absent in both H. bulgarica bulgarica and H. bulgarica canadensis. Additionally, nepalensis has ferwer teeth on the unci than either of the other subspecies, and in nepalensis there is a distinct differentiation into two larger and two or three smaller teeth (Daems and Dumont 1974, Fig.7a-f). (ref. ID; 5932)
Type materials
Holotype: Undissected female mounted in glycerine on a slide, deposited at the KBIN/IRSNB. (ref. ID; 5932)
Paratypes: Ten slides of whole animals, five mastax preparations, and about 70 specimens preserved in a formaldehyde-glycerine solution, all retained at the Laboratorium voor Dierkunde, Rijksuniversiteit Gent, Belgium. (ref. ID; 5932)
Material examined
About 100 partheogenetic females from Gokyo Tso, east Nepal, 4-5 May 1973, H.J. Dumont leg. (ref. ID; 5932)
Dental formula 7/7. Stylate appendages, sabreshaped cuticular appendage on the trunk and lower lip lacking. (ref. ID; 3505)
Body posteriorly rounded, without caudal appendage. Ventral appendage (limb) as long as body. Brackish-water species, in eastern Baltic. (ref. ID; 4596)
The animals examined are morphologically the same as typical H. fennica in all points. They lack a ventro-anterior lip and caudal appendages, the spine formula of the ventral arm is 4/4, and it has nine terminal filaments. Filaments on the other arms are: dorsal, eight; ventrolateral, eight; dorsolateral, seven. The trophi are typical as well. The uncus has eight teeth the upper limit known for this species. The manubria are strongly built and block shaped. The only point on which the Waldsea specimens differ from material from other localities in size: total body length is 360 to 420 µm, which is exceptionally long. (ref. ID; 5932)
SEM micrograph illustrating trophi. (ref. ID; 6844)
Material examined
A large number of parthenogenetic females from Waldsea Lake, Saskatchewan, Canada, 23 July 1974, qualitative plankton haul from 9 m to surface, U.T. Hammer leg. (ref. ID; 5932)
Measurements
Length of the trunk 125-150; height of trunk 155; length of the ventral arm 125; length of the bifurcate filamentous appendages on this arm 100; length of the dorsal arm 85-90; length of its bifurcate filamentous appendages 40-80 µm. (ref. ID; 3505)
Length of body 120-300 µm. (ref. ID; 4596)
Body conical, fairly large. Ventral arm has three pairs of spines and eight filaments. Trophi characteristic with 5 teeth on either side. (ref. ID; 2715)
H. intermedia and H. mira have been carefully studied by Hauer (1941), and their variability has been explored in full detail. They have a tendency to cooccur (Hauer 1941) without overlap in morphological features or intermediate forms, a strong argument in favour of their status as food species. A strongly developed lower lip in H. mira is lacking in H. intermedia, and the teeth formula on the unci is equally different: 3.3/3.3 in mira, 3.2/3.2 in intermedia. An excellent distinctive character at lower magnification is the structure of the ventral arm, well corresponding to Hauer's (1941) figures: three pairs of strong hooks in mira, only one in intermedia. (ref. ID; 2825)
Body conical, fairly large. Ventral arm with 3 pairs of spines and 8 filaments. (ref. ID; 3180)
Measurements
Overall length 350 µm. (ref. ID; 2284)
Total length 220; ventral arm 160 µm. (ref. ID; 2715)
Body large, conical. Ventral arm with three pairs of spines and 8 filaments. (ref. ID; 1804)
Body large, of the same shape as that of H. intermedia. Ventral arm has three spines and 8 filaments. The trophi has 6 teeth on either side. (ref. ID; 2715)
The ventro-anterior lip and cylindrical caudal appendages are well developed. The unci have six teeth. The ventral arm has three pairs of spines plus eight filaments on four sockets. The dorsal arm has six filaments, and the ventrolateral and dorsolateral arms have nine filaments each. The body is conical in shape and 190-240 µm long. There are no spines on the dorsolateral arm. (ref. ID; 5932)
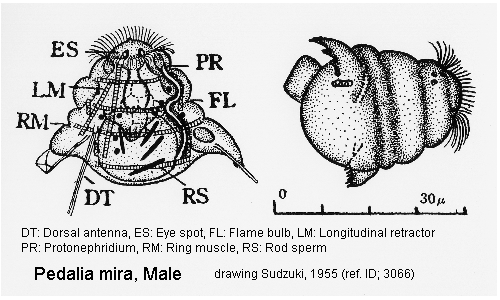
Male: Male present. (ref. ID; 3066)
Quote from ref. ID; 3066
Measurements
Total length 285 µm. (ref. ID; 1804)
Total length 300; resting egg 112 µm. (ref. ID; 2715)
The Katrine Lake animals have a conical body without cylindrical caudal appendages; length 120 to 165 µm. A ventro-anterior lip is present, but it is less apparent in some 1977 specimens collected than in those from 1973. The ventral arm is as long as the body. Its spine formula is 4/4 (four spines on each side), and it has eight terminal filaments implanted on four stems. The dorsal arm has seven terminal filaments implanted on three stems, the middle one bearing three filaments, a character consistent in all specimens examined. The ventrolateral arm has nine filaments. In the plane of the unci, the trophi have crescent-shaped manubria, divided into five parts of which only three are sometimes clearly visible, and the tooth formula on the unci is 11/11. The crescent-shaped manubria in specimens examined here are typical for this species and correspond to the figure given by Hauer (1957), although he did not draw special attention to this feature. The Pond WF7 specimens are similar in most respects to those from Katrine Lake. However, the ventral arm has nine or ten terminal filaments on four or five stems and the tooth formula on the unci is usually 10/10 (ocassionally 10/11 or 11/11). (ref. ID; 5932)
Remarks
The animals from Katrine Lake and from Pond WF7 are morphologically very close to Pedalia fennica var. polyodonta Hauer (Hauer, 1957), now considered, correctly in our opinion, to be a valid species (Kutikova 1970). The most important difference is that Hexarthra polyodonta polyodonta lacks a ventral-anterior lip, whereas H. polyodonta jasperina has a distinct lip. The other differences that we noted were that the dorsal arm on polyodonta has usually six and rarely seven filaments, whereas it has seven in all specimens of jasperina examined. The number of filaments on the ventral arm in polyodonta is eight, whereas in jasperina it varies from eight to ten. Although Hauer (1941) doubts the taxonomic value of the presence or absence of the ventro-anterior lip, noting that is may be difficult to see in preserved specimens, we have found that with proper specimen orientation, the lip when present can easily be seen. As a result of our examination of Hexarthra spp. from various parts of the world, we consider that this lip character may be of subspecific value, because geographically isolated populations which are identical in many basic characters (e.g. structure of trophi, caudal appendages, spines on dorsolateral arm) often differ from one another only in the presence or absence of this lip (a point noted agin in the following sections). Here, H. polyodonta jasperina is such a case. Furthermore, it differs ecologically from the nominate subspecies in that total salinity at the type locality of H. polyodonta polyodonta (Lake Van, eastern Anatolia, Turkey) is 22,000 mg/l (Livingstone 1963), due mainly to NaCl (pH 9.6). Although the salinity of Katrine Lake is high in comparison with all other waters within 200 km (Anderson 1974), it is low in comparison with Lake Van. (ref. ID; 5932)
Type material
Holotype: Undissected female mounted in glycerine on a slide has been deposited at the KBIN/IRSNB (Institut Royal des Sciences naturelles de Belgique, Brussels). (ref. ID; 5932)
Paratypes: Ten slides of whole animals, five mastax preparations, and about 40 specimens preserved in a fromal-dehyde-glycerine solution. Half the paratype material is in the Laboratorium voor Dierkunde, Rijksuniversiteit Gent, Belgium, and half in the Canadian Wildlife Service laboratory, Department of Biology, University of Calgary, Canada. (ref. ID; 5932)
Material examined
About 70 specimens, all parthenogenetic females, from Katrine Lake, Alberta, Canada, 11 July 1973, R.S. Anderson leg., and from pond WF7 near Camrose, Alberta, Canada, 24 August 1976, D.J. Robinson leg. (ref. ID; 5932)