Oxytricha
Oxytricha Bory de St. Vincent, 1824 (ref. ID; 2857) or 1825 (ref. ID; 662, 2014), Ehrenberg, 1831 (ref. ID; 4697) or 1839 (ref. ID; 4905), Bory de Saint-Vincent in Lamouroux, Bory de Saint-Vincent & Deslongchamps, 1824 (ref. ID; 4894)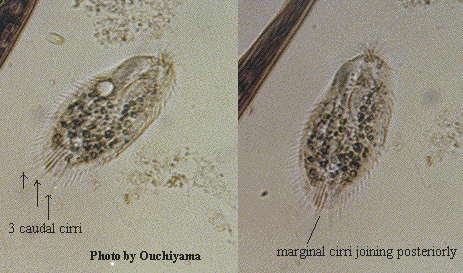
Class Polyhymenophora: Order Hypotrichida: Family Oxytrichidae (ref. ID; 4701)
Family Oxytrichidae Ehrenberg, 1838: Subfamily Oxytrichinae Jankowski, 1979 (ref. ID; 4894) Synonym Histrio Sterki (ref. ID; 1618); Opisthotricha Kent (ref. ID; 1618); Oxitricha Bory, 1824 (ref. ID; 4894) or 1826 (ref. ID; 2014); Steinia Diesing (ref. ID; 1618) [ref. ID; 2014]
Highly flexible, ellipsoidal body with AZM a third to quarter the body length. 2 rows of marginal cirri joining posteriorly serving to distinguish it from the related Opisthotricha and Tachysoma. With or without caudal cirri. Typical arrangement of front-ventral cirri and transverse cirri present. The 3 most anterior cirri large and strong. 2 macronuclei, each with an adjacent micronucleus. Many species have been described. This is the most species-rich genus in the hypotrichs. Kahl's (1932) 7 subgenera are now recognized as genera, however, Jankowski (1979) returns to the amended subgenera and adds 2 new ones.
Quote; Colin R. Curds, Michael A. Gates and David McL. Roberts "British and other freshwater ciliated protozoa Part II Ciliophora: Oligohymenophora and Polyhymenophora" Cambridge University Press, 1983 (ref. ID; 2014) [ref. ID; 2857]
Species of these genera Oxytricha, Sterkiella, Stylonychia, and Histriculus frequently look alike to untrained workers. Thus, they have often been confused. Berger & Foissner (1997) solved puzzle by using morphological and ontogenetic traits, which clearly these and other oxytrichid genera from each other. Briefly, Oxytricha is distinctly different from the other genera by its morphogenetically active postoral cirrus V/3 (Berger & Foissner 1997) and the small subunit ribosomal RNA gene sequences (Schlegel et al. 1991). Thus, it belongs to the subfamily Oxytrichinae, whereas the other genera, in which cirrus V/3 does not participate in anlagen formation, belong to the Stylonychinae. Within this group, only Histriculus lacks caudal cirri and has confluent marginal cirral rows, clearly separating it from Sterkiella and Stylonychia. The latter genera differ morphologically, at the present state of knowledge, mainly in the arrangement of the undulating membranes (intersecting in Sterkiella, parallel in Stylonychia) and the buccal field (narrow in Sterkiella, rather broad-triangular in Stylonychia). The ontogenetic difference noted by Berger and Foissner (1997), namely, that cirral streaks V and VI of the opisthe originate de novo, holds only for Sterkiella cavicola, type of the genus. In S. histriomuscorum (Oxytricha trifallax) and S. nova, these anlagen are generated by cirrus V/4, as in Stylonychia spp. (Wirnsberger et al. 1985, 1986). However, Sterkiella nova and Stylonychia mytilus (type of the genus) differ distinctly in certain gene sequences (Hoffman & Prescott, 1997) and the allozyme pattern (Schlegel 1985; Schlegel and Steinbruck 1986), that is, are distinct genera, in spite of the rather inconspicuous morphological and ontogenetic differences. (ref. ID; 2857) [ref. ID; 4894]
Improved characterization; Adoral zone of membranelles formed like a question mark. Undulating membranes in Oxytricha pattern. Frontoventral cirri in V-shaped pattern. Postoral ventral cirri in dense cluster underneath buccal vertex. Two pretransverse ventral and five, rarely four transverse cirri. One right and one left row of marginal cirri. Usually five or six, rarely four dorsal kineties. Caudal cirri present. Primordia V and VI of the proter originate from cirrus V/4 and V/3, respectively. Dorsal morphogenesis in Oxytricha or Urosomoida pattern. (ref. ID; 4894)
Remarks; Bory de Saint-Vincent in Lamouroux et al. (1824) used the spelling Oxitricha when establishing the genus. The first reviser (Ehrenberg 1838) changed it to Oxytricha, subsequently accepted by all revisers (Kahl 1932; Borror 1972; Corliss 1979). For a detailed discussion of the rather complicated type species problem, see Foissner (1989). Oxytricha is the most voluminous genus within the oxytrichids, comprising about 50 valid species and about 60 species indeterminata (H. B., unpubli. data). Only a quarter of the valid species has been described in detail. Actinotricha is a little known genus which has been synonymized with Tachysoma (Borror 1972) and Oxytricha (Song et al. 1991). Very likely, it is closely related to the O. setigera group because the frontoventral cirri are distinctly displaced posteriorly. Opisthotricha was established for Oxytricha species with long caudal cirri (Kent 1880-1882). However, this feature is valuable for species distinction only. Non-specialists easily confuse Oxytricha with the stylonychid genera Sterkiella (body only slightly flexible, posterior postoral ventral cirrus V/3 distinctly separated from cirri V/4 and IV/2), Histriculus (body very rigid, confluent marginal rows, no caudal cirri) and Stylonychia (body very rigid, undulating membranes side by side) where cirrus V/3 is, however, not involved in primordia formation. (ref. ID; 4894)
Type species (subsequent designation by Foissner 1989); Oxytricha granulifera Foissner & Adam, 1983 (ref. ID; 4894)
- Oxytricha aeruginosa Wrzesniowski, 1870 (ref. ID; 1621) reported year? (ref. ID; 5462) or Wrzesniowskiego, 1866 (ref. ID; 4519) reported author and year? (ref. ID; 191)
- Oxytricha affine Stein, 1859
See; Trachelostyla affine (ref. ID; 2365) - Oxytricha affinis Stein, 1859
See; Gonostomum affine (ref. ID; 2294) - Oxytricha agillis (ref. ID; 191)
- Oxytricha alba Fromental, 1876
See; Holosticha pullaster (ref. ID; 4609) - Oxytricha alfredkahli (ref. ID; 4730)
- Oxytricha auricularis Claparede & Lachmann, 1858
See; Epiclintes ambiguus (ref. ID; 1621) - Oxytricha balladyna Song & Wilbert, 1989 (ref. ID; 4609)
See; Oxytricha opisthomuscorum (ref. ID; 2846) - Oxytricha bifaria Stokes, 1887 (ref. ID; 1308, 1621, 7565) reported year? (ref. ID; 1618, 3342) reported author and year? (ref. ID; 191, 4471, 7251, 7608, 7717, 7721)
- Oxytricha bimembranata Shibuya, 1929 (ref. ID; 1621)
- Oxytricha buitkampi Dragesco & Dragesco-Kerneis, 1986 (ref. ID; 4730)
- Oxytricha candens (Kahl) (ref. ID; 5462)
Syn; Steinia candens Kahl (ref. ID; 5462) - Oxytricha cavicola Kahl, 1935 (ref. ID; 2857) reported author and year? (ref. ID; 1620)
See; Sterkiella cavicola (ref. ID; 2857) - Oxytricha chlorelligera Kahl, 1932 (ref. ID; 4488, 4609) reported author and year? (ref. ID; 1621, 1629)
- Oxytricha crassistilata (Kahl, 1932) Borror, 1972 (ref. ID; 2846) reported author and year? (ref. ID; 191)
See; Oxytricha opisthomuscorum (ref. ID; 2846) - Oxytricha crassistillata Hemberger, 1982 (ref. ID; 4719)
- Oxytricha discifera Kahl (ref. ID; 1621, 2117)
- Oxytricha dubia O.F. Muller (ref. ID; 2117)
- Oxytricha echinata Claparede & Lachmann, 1859
See; Oxytricha pellionella (ref. ID; 3116), Tachysoma pellionella (ref. ID; 1621) - Oxytricha enigmatica Dragesco & Dragesco-Kerneis, 1986
See; Hemigastrostyla enigmatica (ref. ID; 4905) - Oxytricha fallax Stein, 1859 (ref. ID; 1308, 1621, 1629, 2245, 4488, 4609) reported year? (ref. ID; 1219, 1618, 3698, 4111) reported author and year? (ref. ID; 191, 3736, 4471)
- Oxytricha fennica Hemberger, 1982 (ref. ID; 4719) reported author and year? (ref. ID; 191)
- Oxytricha ferruginea Stein, 1859 (ref. ID; 4519, 4609)
- Oxytricha fromenteli (ref. ID; 4730)
- Oxytricha furcata Smith, 1897 (ref. ID; 1621)
- Oxytricha furruginea Stiller (ref. ID; 1629, 2245)
- Oxytricha fusca Perty, 1852
See; Urostyla grandis (ref. ID; 1621) - Oxytricha fusiformis (ref. ID; 191)
- Oxytricha germanica (ref. ID; 4730)
- Oxytricha gigantea Horvath, 1933 (ref. ID; 4751) reported author and year? (ref. ID; 191)
- Oxytricha granulifera Foissner & Adam, 1983 (ref. ID; 2128, 2308, 4719) reported author and year? (ref. ID; 191, 4471)
- Oxytricha haematoplasma Blatterer & Foissner, 1990 (ref. ID; 4609, 4781 original paper) reported author and year? (ref. ID; 1629)
- Oxytricha hymenostoma Stokes, 1887 (ref. ID; 1621, 1629, 4609) reported author and year? (ref. ID; 3821)
- Oxytricha islandica Berger & Foissner, 1987 (ref. ID; 2308)
- Oxytricha kessleri Wrzesniowski, 1877
See; Holosticha kessleri (ref. ID; 4609) - Oxytricha lanceolata Shibuya, 1930 (ref. ID; 1620, 2128)
- Oxytricha longa Hemberger, 1982 (ref. ID; 4719) reported author and year? (ref. ID; 191)
- Oxytricha longicaudata Wright, 1862
See; Mitra retractilis (ref. ID; 1621) - Oxytricha longigranulosa Berger & Foissner, 1989 (ref. ID; 2308) reported author and year? (ref. ID; 4471)
- Oxytricha ludibunda Stokes, 1891 (ref. ID; 1308, 1621) reported year? (ref. ID; 1618)
- Oxytricha marina (ref. ID; 1621)
- Oxytricha minor Kahl (ref. ID; 3848) reported author and year? (ref. ID; 1621)
- Oxytricha muscorum Kahl, 1932 (ref. ID; 4695) reported year? (ref. ID; 3698)
See; Cyrtohymena muscorum Kahl, 1932 (ref. ID; 4894) - Oxytricha mystacea Stein, 1859
See; Gastrostyla mystacea (ref. ID; 4609) - Oxytricha nova Klobutcher et al., 1981 (ref. ID; 2857) reported author and year? (ref. ID; 4471)
See; Sterkiella nova (ref. ID; 2857) - Oxytricha opisthomuscorum Foissner et al, 1991 (ref. ID; 2128, 2846)
- Oxytricha ottowi Foissner (ref. ID; 2308 original paper)
- Oxytricha ovalis (ref. ID; 1621)
- Oxytricha ovata (Kahl) (ref. ID; 5462)
Syn; Opisthotrica ovata Kahl (ref. ID; 5462); Steinia sapropelica Kahl (ref. ID; 5462) - Oxytricha pellionella O.F. Muller (ref. ID; 1621) or (O.F. Muller) Stein, 1859 (ref. ID; 3116)
Syn; Oxytricha echinata (ref. ID; 3116); Tachysoma agilis Stokes, 1887 (ref. ID; 3116); Tachysoma pellionella (ref. ID; 1621, 3116) - Oxytricha planctontica (Horvath, 1933) (ref. ID; 4701)
- Oxytricha platystoma Ehrenberg, 1831 (ref. ID; 4609) reported year? (ref. ID; 1335)
See; Steinia platystoma (ref. ID; 4609) - Oxytricha proximata Shibuya, 1930 (ref. ID; 1620)
- Oxytricha pseudosimilis Hemberger, 1982 (ref. ID; 4719) or 1985 (ref. ID; 4697 original paper) reported author and year? (ref. ID; 191)
- Oxytricha rubra Buitkamp (ref. ID; 2365 original paper)
- Oxytricha rubra Ehrenberg, 1838
See; Keronopsis rubra - Oxytricha saltans (Cohn, 1866) (ref. ID; 4905)
Syn; Actinotricha hyalina Perejaslawzewa, 1885 (ref. ID; 4905) - Oxytricha saprobia Kahl, 1932 (ref. ID; 4609) or 1935 (ref. ID; 3116) reported year? (ref. ID; 1219, 1621, 1629)
- Oxytricha selvatica Hemberger, 1985 (ref. ID; 4697 original paper)
- Oxytricha seseris (ref. ID; 662)
- Oxytricha setigera Stokes, 1891 (ref. ID; 662, 1308, 1621, 1629, 3116, 4609) reported year? (ref. ID; 1618, 4842)
See; Oxytricha opisthomuscorum (ref. ID; 2846) - Oxytricha similis Engelmann, 1862 (ref. ID; 4488, 4609) reported year? (ref. ID; 5462) reported author and year? (ref. ID; 1629)
Syn; Opisthotrica similis Engelmann (ref. ID; 5462) - Oxytricha siseris Vuxanovici, 1963 (ref. ID; 662)
- Oxytricha sphagni (Kahl, 1932) Borror, 1972
See; Oxytricha opisthomuscorum (ref. ID; 2846) - Oxytricha stenocephala Borror, 1963
See; Hemigastrostyla stenocephala (ref. ID; 4905) - Oxytricha strenua Engelmann, 1862
See; Gonostomum strenua (ref. ID; 2294) - Oxytricha terrestris Hemberger, 1982 (ref. ID; 4719) reported author and year? (ref. ID; 191)
- Oxytricha tricornis Milne, 1886 (ref. ID; 1621)
- Oxytricha trifallax (ref. ID; 4471, 4969)
See; Sterkiella histriomuscorum (ref. ID; 2857) - Oxytricha tubicola Gruber, 1877 (ref. ID; 1621)
- Oxytricha wrzesniowskii Mereschkowsky
See; Holosticha kessleri (ref. ID; 4609)
Oxytricha bifaria Stokes, 1887 (ref. ID; 1308, 1621, 7565) reported year? (ref. ID; 1618, 3342) reported author and year? (ref. ID; 191, 4471, 7251, 7608, 7717, 7721)
Descriptions
Right side convex; left side flattened; posterior end pointed; fresh water infusion. (ref. ID; 1618) [ref. ID; 7608]Doublets (Strain C9): Strain C9 of Oxytricha bifaria is about 100 um long. The doublets are characterized by a slightly shorter body. Doublets show homopolar organization with two functional peristomes anteriorly. Five morphological states of doublets may be described. (ref. ID; 7608)
- Type 1. The type 1 consists of two single hemicells fused by their longitudinal mid-dorsal surfaces: their spatial arrangement leads their longitudinal axes to coincide and their ventral surface to bend longitudinally, producing a sort of groove, so that the transverse section assumes a four-tip-star form.
- Type 2. The type 2 is only apparently similar to type 1: the longitudinal dorsal line uniting the two singlets is a paramedial one so that the two longitudinal axes, although still parallel, do not coincide any more and both the anterior and the posterior ends of the two halves of the doublet-2 are separated from each other. The ventral surfaces are not bent any more and look quite normal: this observation accounts for the transverse section of this kind of doublet, which is lozenged.
- Type 3. The type 3 represents a kind of doublet somehow peculiar, in comparison with both type 1 and type 2, because of the relative spatial disposition of the two cellular components: the two anterior ends are clearly separated from each other, being the two hemicells united longitudinally by their dorsal surfaces, but only at the level of the posterior half. Their longitudinal axes not only do not coincide any more (cfr. type 1) but also start diverging at the anterior ends of the cells. (This seems to suggest that the anterior regions are less stable when fused that the posterior ones).
- Type 4. The type 4 shows a very distinct shape, because of the divergence between the two halves of the doublet, now evident beyond any doubt. Moreover the two hemicells bend ventrally at the level of their middle part, so that actually they form two different angles; the angle (alpha) between the posterior halves of the hemicells roughly ranges around 50 degrees, while the angle (beta) between their anterior parts is of about 120 degrees. The two cell bodies are now well individuated, although a large area of their dorsal membrane still unites them as if stretched by their pulling apart.
- Type 5. The type 5 doublet in a sense is not a doublet s.s. any more: the two oxytrichas indeed are now separated almost completely and have an almost normal shape: however their posterior ends, now subconical, are still connected by a sort of cytoplasmic "stalk", which represents what remains of the initial 100% longitudinal union. The life of the type 5 is very short, never lasting more than 30 min. (ref. ID; 7608)
Cyst:
- I. The young cysts.
The cyst wall. The first observable modification which was reported for an encysting O. bifaria (Ricci et al. in press) was the production of an external jelly coat tightly adhering to the cell surface. By electron microscopy, such a jelly coat proved to consist of lamellar material which at first is irregularly arranged all around the spherical cell while successively it becomes more and more regularly patterned along the cell surfaces contour, giving rise to the ectocyst. An empty pericellular space appears between the cell and this external lamellar layer; it increases progressively and rather regularly up to 3 um in thickness within 1-2 hours. It must be recalled that during this phase the contractile vacuole is still working and it will go possible to state whether and how its activity could be related to the widening of the pricellular space and to the progressive reduction of the cell volume. Then a double coat become visible above the cell surface. The outer one is very homogeneous, slightly electrondense, constant in thickness (C ~0.5 um) and regular in shape; the second occupies the irregular space between the first one and the cell surface, now clearly wrinkled. The remaining pericellular space underneath the outer lamellar layer (ectocyst), is progressively filled (starting from its inner part) by the mesocyst, made by many layers of fibers which are oriented perpendicularly to each other. One day-old cysts show the classical four layers configuration of the wall, already described for several other species: i) the ectocyst, namely the external lamellar layer now well organized into an internal subspherical part and into an external one which forms the finger-like protrusions (flp); ii) the mesocyst, about 2 um thick, structured into many concentric layers; iii) the endocyst very homogeneous and regular both in shape and thickness (0.3-0.5 um); iiii) the granular layer, the most internal one regularly discontinuous and uniting the spherical cyst wall to the wrinkled cell surface.
The cytoplasm and the nucleus. During the first phases of encystment when only the outer lamellar layer of the cyst wall is formed, orderly arrays of rough endoplasmic reticulum are prominent somehow surrounding other cellular organelles. The precursors of the cyst wall are evident as densely packed discs, contained within elongated, circa radially oriented vesicles, at least two different kinds of vesicles can be distinguished. Their distribution suggests that they move from deeply in the cytoplasm when they are probably synthesized, to the pericellular space. The subcortical area shows many flattened cysternae, which later disappear when the surface assumes a regular wrinkled geometric pattern buttressed by microtubules similarly oriented. At the cyst completion the rough endoplasmic reticulum is not evident any more and many rounded small vesicles containing light electrondense material appear underneath the now wrinkled cell surface. The appearance of these vesicles in this particular stage would seem to suggest their involvement in the process or release of cyst components. The resorbtion of ciliary organelles is not yet complete, the contractile vacuole is still functioning, and mitochondria, paraglycogen granules and lipid droplets are scattered throughout the cytoplasm. No appreciable chage is noticeable in the macronuclear chromatin arragement. (ref. ID; 7717) - II. The resting cysts.
The cyst wall. The 14 day-old cysts loose their roundish shape and appear elongated and compressed in some points. The lamellae of the ectocyst run parallely to each other and the more external ones rise at regular distances, to form the typical flp among which bacteria are often trapped. The mesocyst consists of two layers, ecto- and endo-mesocyst which can be either separated by a less condensed and rather discontinuous zone or not. However, which ever the ultrastructure of the mesocyst, a lot of heavily electrondense granules are scattered amid the fibres of the ectomesocyst; these granules are regularly arranged into a compact pattern just underneath the ectocyst. The endocyst appears unmodified while the granular layer shows a coarser and more electrondense granulation than in the early cyst stages. Seldom some cysts have been found without the endocyst and with a cytoplasm heavily vacuolized.
The cytoplasm and the nucleus. Many major traits must be recalled: a) the ciliary organelles are completely resorbed, b) the subcortical microtubules are still present; c) the cytoplasm is very condensed; d) the mitochondria are grouped in clusters; e) the paraglycogen granules are localized mainly at one pole; f) the autophagic vacuoles are rare. The macronuclear chromatin appears to be arranged in spherical bodies regularly scattered throughout the nuclear matrix. (ref. ID; 7717) - III. The young empty cysts. A certain percentage of encysting oxytrichas proves to be unable to complete their cyst wall and, consequently, to survive, so that a certain amount of young empty cysts is usually found amid the others, physiologically perfect. The electron microscopical observation shows that their wall is not well differentiated and consists of only an amorphous mesocyst and of a typical ectocyst which, however, is broken in some points. Many bacteria passing through these interruptions of the ectocyst penetrate the cyst wall and reach the cell surface. The cytoplasm is very rarefied but in some cases cyst wall precursors are still present. (ref. ID; 7717)
Remarks
Oxytricha bifaria is a freshwater hypotrich; it is subelliptical in shape (110-120 by 60-70 um) and lives by feeding on bacteria and dividing vegetatively. Under particular internal and (or) external conditions, however, cultures can undergo any four different fates; the cells can form pairs (Ricci 1981), cysts (Ricci et al. 1985); giants (Ricci and Riggio 1984), or doublets (Banchetti and Ricci 1986). The giants were described by Dawson as early as 1919, and only very recently Ricci and Riggio (1984) investigated their differentiation. Once a giant is isolated in bacterial medium without any Oxytricha as food, it returns progressively to its normal shape and dimensions through a series of three successive cell divisions (Ricci and Riggio 1984). (ref. ID; 7251)Measurements
About 250 um long. (ref. ID; 1618)140-180x70-120 um. (ref. ID; 3342)
Oxytricha fallax Stein, 1859 (ref. ID; 1308, 1621, 1629, 2245, 4488, 4609) reported year? (ref. ID; 1219, 1618, 3698, 4111) reported author and year? (ref. ID; 191, 3736, 4471)
Descriptions
Body outline ellipsoid, posterior region broadly rounded; flexible; dorsal surface convex; ventral side flattened and equipped with cirri (frontal 8, ventral 5, transversal 5); right and left marginal cirri near meet as a continuos row posteriorly (Stylonychia, with marginal cirri interrupted at posterior end, with long caudal cirri); buccal area with well-developed adoral zone of membranelles and a long undulating membrane at the right side; cytostome located at the posterior end of the buccal area; 1 contractile vacuole near to the left margin in the vicinity of the posterior end of the buccal area. (ref. ID; 1219)Measurements
Length about 150 um. (ref. ID; 1219, 1618)Oxytricha gigantea Horvath, 1933 (ref. ID; 4751) reported author and year? (ref. ID; 191)
Descriptions
The morphogenesis begins apokinetally with the proliferation of basal bodies in the area between the postoral ventral cirri and transverse cirri. The replication bands have moved about 2/3 of the length of the macronuclear segments in this stage. A long oral primordium is formed. Its narrowed anterior part elongates to the level of the cytostome and splits up in the middle region to form skreaks I and II. The postoral ventral cirri V/3 and V/4 are modified to primordia. Next, the postoral ventral cirrus IV/2 is disorganized. Five streaks of the opisthe are recognizable at this stage. At the same time, the buccal cirrus (II/2) and the cirrus III/2 organize primordia for the proter. Three other primordia are present at the level of the cytostome. At least the middle and the right primordia are deribed from a streak of the opisthe. Six fronto-ventral-transverse primordia are discernible in the next stage in both the proter and the opithe. Cirrus IV/3 has been resorbed. Some loosely arranged basal bodies are present between the right streak (primordium VI) of the proter and the primordium IV of the opisthe. Morphogenesis continues with the elongation of the fronto-ventral-transverse streaks and the formation of marginal primordia in the anterior and middle part of the right marginal row. The proliferation of new basal body pairs occurs at 2 levels in the dorsal kineties 1, 2, and 3. The parental membranes commence with reorganization when the undulating membranes of the opisthe are formed. The anterior cirri are already segregated from the fronto-ventral-transverse streaks. Both the right and left marginal primordia are elongated. The primordia of the dorsal kineties 4 originate at the anterior end of the right marginal primordia at about the time when the macronuclear segments are fusing. The rest of the morphogenetic events is very similar to that known of other oxytrichids with 18 frontal, ventral, and transverse cirri, e.g., Oxytricha granulifea and Stylonychia spp. The constant number of 18 cirri is formed within the fronto-ventral-transverse primordia I-VI as follows: 1, 3, 3, 3, 4, and 4 cirri. (ref. ID; 4751)Comments
Kahl (1935) transferred O. gigantea to the genus Urosoma Kowalewski, 1882, because of its elongated and posteriorly converging body shape. The confused history of the genus Urosoma is explained by Foissner (1983). When O. gigantea was redescribed by Berger and Foissner (1987) as U. gigantea (Horvath, 1933) Kahl, 1935, the morphogenesis was still unstudied. Likewise, the morphogenesis of the type-species of Urosoma, U. cienkowski, is unknown, so that the decision to which genus Horvath's species belongs cannot be clarified unequivocally. Foissner (1983), however, investigated the division of the closely related species U. macrostyla. A comparison of the morphogenesis of these species argues for the retention of O. gigantea in the genus Oxytricha. The main difference between the morphogenetic patterns exists in the origin and formation of the fronto-ventral-transverse streaks. "Primary streaks" are formed in U. macrostyla. They divide in the middle stages of the morphogenesis and form the "secondary primordia" that are the primordia for the infraciliature of the proter and the opisthe. This type of morphogenesis occurs also in G. affine and Tachysoma pellionella. In contrast, in O. gigantea only streaks IV (?), V, and VI of the proter arise from streak IV of the opisthe. This type of fronto-ventral-transverse streak formation appears also in e.g., O. granulifera and Urosomoida agiliformis. The second character that supports the inclusion in the genus Oxytricha is the position of the cirrus III/2, that is the cirrus behind the right frontal cirrus. Already Foissner (1982) recognized that in typical members of the genus Urosoma this cirrus is always situated anteriorly to the cirrus VI/4. Thus, the cirri III/2, VI/4, VI/3, and IV/3 form nearly a line. On the other hand, in O. gigantea and other Oxytricha-species, e.g., O. granulifera and O. lanceolata the cirrus III/2 is situated left of the area between the cirri VI/4 and VI/3. Thus, these cirri form a V-like figure. Very probably, the position of the cirrus III/2 is the most appropriate character with which to separate Oxytricha and Urosoma, since Urosoma-species which do not have the tail-like posterior end of the type-species, U. cienkowski are known to exist. Borror (1972) and Hemberger (1982) synonymized O. gigantea Horvath, 1933 with Urosomoida agilis (Engelmann, 1862) Hemberger, 1985 and O. parallela Engelmann, 1862, respectively. However, it is clear from the original description and the redescription of Berger and Foissner (1987) that it can be separated from these and other related species by the characters body size, morphogenetic pattern, subpellicular granules, and the habitat. (ref. ID; 4751)Oxytricha hymenostoma Stokes, 1887 (ref. ID; 1621, 1629, 4609) reported author and year? (ref. ID; 3821)
Descriptions
Oxytricha hymenostoma has a subelliptical body, measuring ~100x40 um. There are 2 ovoid macronuclei (rarely, 3) micronuclei, lying on the left side of a macronuclear pocket. Our determination was based on the presence of 2 undulating membranes, as described by Stokes. (ref. ID; 3821)Oxytricha lanceolata Shibuya, 1930 (ref. ID; 1620, 2128)
Descriptions
Moss and soil inhabiting species. O. lanceolata can be characterized as follows: length in vivo 80-120 um (75-110 um in protargol, slides), 2 macronuclear nodules, 2 micronuclei, 23-30 (M=27-28) adoral membranelles, 25-33 (M=28-29) right marginal cirri, 25-35 (M=31-32) left marginal cirri, 5 transverse cirri, 3 caudal cirri, 4 dorsal kineties. The most important character, as compared with the typical oxytrichid ventral cirral pattern and number, is the reduced number of dorsal kineties with kinety 4 distinctly shortened posteriorly. Furthermore, all populations lack special cortical granules, which greatly facilitates in vivo separation from an other frequent and rather similar soil species, viz. O. granulifera. (ref. ID; 2128)Oxytricha ludibunda Stokes, 1891 (ref. ID; 1308, 1621) reported year? (ref. ID; 1618)
Descriptions
Ellipsoid; flexible; fresh water among sphagnum. (ref. ID; 1618)Measurements
100 um long. (ref. ID; 1618)Oxytricha muscorum Kahl, 1932 (ref. ID; 4695) reported year? (ref. ID; 3698)
See
Cyrtohymena muscorum Kahl, 1932 (ref. ID; 4894)Descriptions
The paroral formation is divided into two parts (PF1 and PF2). PF1, the most anterior of the two, measures 22.2-24.3 um in length and is composed of 2 adjoining and parallel rows of 64-66 kinetosomes each. PF2 has a length of 25.5-26.4 um and is composed of two different areas:- a) an anterior area, of 16.5-18.3 um in length, composed of rows with 46-48 kinetosomes. These rows are arranged with an inclination of approximately 45 degrees with respect to the axis of PF2 and their kinetosomes have derivates situated between 2 contiguous rows, perpendicularly to the axis of these rows. The ensemble therefore has the appearance of a gird. (ref. ID; 4695)
- b) a posterior area, of 7.5-8.2 um in length, composed of 2 adjoining and parallel rows of 20-22 kinetosomes each. (ref. ID; 4695)