Amphileptus
Amphileptus Ehrenberg, 1830 (ref. ID; 2013)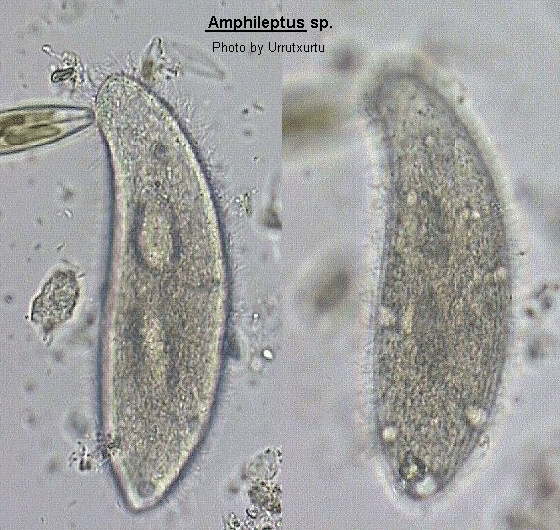
Body laterally compressed, highly elongate with anterior neck-like region which bends towards the dorsal edge. Oral aperture a slit on convex edge of neck region, extends less than halfway down the body. Ciliation present on both lateral surface although there is a tendency to some reduction on the left surface resulting in it being difficult to distinguish. Ciliation on right surface is extensive and forms longitudinal rows which converge on each other in the anterior region. There is a distinctive area of cilia along the oral slit forming a mane-like brush. Trichocysts commonly present particularly in neck. Macronucleus in 2 to 4 spherical parts with single micronucleus placed between macronuclei. Many contractile vacuoles present, usually lying along dorsal and ventral edges. The recommendations and finding of Canella (1960) and Frys-Vesavel et al. (1976) have been adopted here so that the genus Hemiophrys has been submerged into the above genus. This is because cilia do appear to be present on the left surface even though difficult to resolve without the use of silver impregnation techniques.
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013)
- Amphileptus affinis (ref. ID; 4612)
- Amphileptus anaticula Claparede & Lachmann, 1859
See; Acineria incurvata Dujardin, 1841 (ref. ID; 1601) - Amphileptus anser Ehrenberg, 1838
See; Litonotus cygnus (O.F. Muller, 1776) Wrzesniowski, 1870 (ref. ID; 1622), Dileptus anser (ref. ID; 3115), Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683) - Amphileptus branchiarum Wenrich (ref. ID; 1618)
- Amphileptus carchesii Stein, 1867 (ref. ID; 1622, 1629, 2245, 2570, 2574, 4612)
Syn; Hemiophrys fossigera Kahl, 1935 (ref. ID; 4612) - Amphileptus carchesii var. major (ref. ID; 4612)
- Amphileptus claparedei Stein, 1867 (ref. ID; 1219, 1335, 1622, 1629, 2245, 2570, 2574, 3115) reported year? (ref. ID; 1618), claparedii Stein, 1867 (ref. ID; 4612)
Syn; Amphileptus meleagris Claparede & Lachmann, 1858 (ref. ID; 1219, 1622, 2245, 3115) or 1861 (ref. ID; 4612) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 3292) - Amphileptus cygnus Claparede & Lachmann, 1859
See; Dileptus anser (ref. ID; 4683) - Amphileptus fasciola Ehrenberg, 1838
See; Litonotus fasciola (ref. ID; 3115) - Amphileptus flexilis (Stokes, 1885) (ref. ID; 1622)
- Amphileptus gutta Cohn, 1886 (ref. ID; 1622)
- Amphileptus gutta Kalmus, 1929 (ref. ID; 1622)
- Amphileptus incurvatus Lepsi, 1926
See; Acineria incurvata (ref. ID; 1601) - Amphileptus margaritifer Ehrenberg, 1833 (ref. ID; 4612) or 1838 (ref. ID; 3115, 4683)
See; Dileptus anser (ref. ID; 3115, 4612), Dileptus margaritifer Ehrenberg, 1838 (ref. ID; 4683) - Amphileptus marinus (Kahl, 1931) Song, Wilbert & Hu, 2004 (ref. ID; 4942 redescribed paper)
Basonym; Lionotus (Hemiophrys) marina Kahl, 1931 (ref. ID; 4942) - Amphileptus meleagris Claparede & Lachmann, 1858 or 1861
See; Amphileptus claparedei (ref. ID; 1219, 1618, 1622, 2245, 3115, 4612) - Amphileptus monilatus Stokes, 1886
See; Monilicaryon monilatus (ref. ID; 4612) - Amphileptus ovum Dujardin, 1841
See; Trachelius ovum (ref. ID; 3115) - Amphileptus parafusidens Song & Wilbert, 1989 (ref. ID; 4942)
- Amphileptus pleurosigma (Stokes, 1884) Foissner, 1984 (ref. ID; 4488, 4612) reported author and year? (ref. ID; 191, 1629)
Syn; Hemiophrys pleurosigma (Stokes, 1884) Kahl, 1931 (ref. ID; 4612); Litonotus pleurosigma Stokes, 1884 (ref. ID; 4612) - Amphileptus plurivacuolata (Foissner, 1978) (ref. ID; 4730)
Syn; Hemiophrys plurivacuolata Foissner, 1978 (ref. ID; 4730) - Amphileptus procerus (Penard, 1922) Song Weibo & Wilbert, 1989 (ref. ID; 4612) reported author and year? (ref. ID; 1629)
Syn; Hemiophrys procera (Penard, 1922) Kahl, 1931 (ref. ID; 4612); Litonotus procerus Penard, 1922 (ref. ID; 4612) - Amphileptus punctatus (Kahl, 1926) Foissner, 1984 (ref. ID; 4612, 4942) reported author and year? (ref. ID; 1629)
Syn; Lionotus punctatus Kahl, 1926 (ref. ID; 4612) - Amphileptus rotundus Maskell, 1887
See; Trachelis ovum (ref. ID; 1622, 3115) - Amphileptus trachelioides (Zacharias, 1893) Kahl, 1930 (ref. ID; 4613)
- Amphileptus tracheloides (Zacharias, 1893) (ref. ID; 1622, 2245, 2568) or (Zacharias, 1893) Maskell (ref. ID; 3115)
- Amphileptus utriculariae Penard, 1922
See; Loxophyllum utriculariae (ref. ID; 4612), Siroloxophyllum utriculariae (ref. ID; 7355) - Amphileptus viridis Ehrenberg, 1833
See; Dileptus viridis (ref. ID; 4730)
Amphileptus branchiarum Wenrich (ref. ID; 1618)
Descriptions
On the integument and gills of frog tadpoles; swimming individuals killed with iodine. (ref. ID; 1618)Measurements
100-135 by 40-60 um. (ref. ID; 1618)Amphileptus claparedei Stein, 1867 (ref. ID; 1219, 1335, 1622, 1629, 1629, 2245, 2570, 2574, 3115) reported year? (ref. ID; 1618), claparedii Stein, 1867 (ref. ID; 4612)
Synonym
Amphileptus meleagris Claparede & Lachmann, 1858 (ref. ID; 1219, 1622, 2245, 3115) or 1861 (ref. ID; 4612) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 3292)Descriptions
Mouth is on the convex (ventral) side, slit-like, clearly visible only when the ciliature is feeding; ciliation uniform and nearly complete but ciliary rows of the left side of the body are nor distinct and somewhat reduced by comparison with those on the right side; numerous contractile vacuoles irregularly distributed along the ventral and dorsal margins; 2 macronuclei; fission takes place with in cysts. (ref. ID; 1219)Slightly flattened; broadly flask-shaped; with bluntly pointed posterior and neck-like anterior end; cytostome about two-fifths from ventral margin; trichocysts indistinct; dorsal ciliary rows also not distinct; contractile vacuoles irregularly distributed; fresh and salt water, on stalks of Zoothamnium, Carchesium, Epistylis, etc. (ref. ID; 1618)
Measurements
Length 120-150 um. (ref. ID; 1219, 1618)Amphileptus marinus (Kahl, 1931) Song, Wilbert & Hu, 2004 (ref. ID; 4942 redescribed paper)
Basionym
Lionotus (Hemiophrys) marina Kahl, 1931 (ref. ID; 4942)This organism was originally described as a new species under the subgenus Hemiophrys by Kahl (1931), Lionotus (Hemiophrys) marina Kahl, 1931. Our current work has confirmed that it is a valid and distinct species with the characteristic pattern of the genus Amphileptus, hence a new combination is made because the genus/subgenus Hemiophrys is a junior synonym of Amphileptus. Since no ciliary information was previously available for this organism, here we supply an improved diagnosis and detailed data on the infraciliature as well as on the morphology of living cells, based on a Chinese population. (ref. ID; 4942)
Revised diagnosis
Large marine Amphileptus 200-450 um long in vivo with a slender lanceolate body, two macronuclear nodules and several micronuclei; 10-12 left and 20-27 right somatic kineties; two perioral kineties extending posteriorly to the extreme end of the cell; several contractile vacuoles positioned ventrally in the posterior half of the cell; extrusomes bar-shaped, c. 12 um long in vivo, densely arranged along the ventral edge. (ref. ID; 4942)Descriptions
Size highly variable, in vivo from 200 to 450 x 50 to 90 um, but mostly around 300-350 um in length. Cells basically slender lanceolate and changeable in shape, i.e. conspicuously flexible and contractile as commonly seen in most other large pleurostomatids. Nevertheless, specimens mostly with a rounded posterior end, sometimes narrowed in the caudal portion (most frequently in small cells) but never tail-like. Margins of main cell portion from parallel (generally when stretched) to considerable convex. Neck about 1/4-1/3 of cell length, often curving highly backwards. Bilaterally about 1:2-3 flattened in body part, hence strongly compressed considering the large size; when crawling on debris, often slightly twisted even folded. Pellicle thin and flexible, which renders body outline sometimes uneven; no conspicuous grooves or ridges on left side ("dorsal side" when viewed in vivo); beneath cell surface, many tiny (<0.8 um across) cortical granules, sparsely distributed between ciliary rows; cilia sparsely distributed on "dorsal" side and c. 2 um long (very difficult to detect in life). Right side densely ciliated, with more or less distinct grooves, in which kineties originate; cilia on this side about 8-10 um long. Cytoplasm colorless to slightly brown-grayish, often with numerous differently sized greasily shining globules, which render the main part of the body more or less opaque (especially at low magnification). Extrusomes bar-shaped, straight and c. 12 um long, nevertheless, inconspicuous in vivo (detectable only under high magnifications), densely arranged along whole oral region as well as the ventral side, scattered in other body portions. Food vacuoles small (<5 um) and low in number (even unrecognizable), containing likely flagellates and diatoms. Several (3-7) contractile vacuoles about 5-20 um across, pulsating infrequently, positioned near the ventral margin along posterior 2/5 to 1/2 of cell length; the largest vacuoles usually at the posterior of the cell, others smaller and unevenly distributed. Two macronuclear nodules, separated in mid-body, basically ellipsoid in shape, in vivo often as two large transparent areas. After protargol impregnation (and after days of culture), however, macronuclei often different in shape and size, sometimes in 3 or 4 semi-detached or linked parts. Nucleoli small, high in number and evenly distributed. Micronuclei several in number (maximum 5 observed), about 4-5 um across, near or adjacent to macronuclear nodules. Generally not sensitive to disturbance. Movement typical of genus, usually gliding on substrate. When swimming, slowly turning around its longitudinal axis for only a short while. (ref. ID; 4942)- Oral structure: Genus specifically, two densely ciliated perioral kineties (PK1, PK2), one each on the right and left of cytostome extending to the posterior end of the cell, 80% of kinety lengths consist of dikinetids. No typical PK3 present as commonly seen in Litonotus or Loxophyllum (Foissner 1984; Song & Wilbert 1989) though always some basal body pairs are observed near PK2. Nematodesmata long and strongly developed, originating from perioral kinetids and extending to about middle of the cell length. (ref. ID; 4942)
- Somatic ciliature: Somatic ciliature genus-specific. All somatic kineties (20-27 excluding the perioral kinety) on right side being relatively densely ciliated, among which the central ones are anteriorly shortened and thus make an anterior suture. On the left side, all kineties of full body length and cilia much more loosely arranged. Dorsal brosse (DK) composed of close-set basal body pairs, terminating about 1/2 of cell length and continuous posteriorly with a row of monokinetids. (ref. ID; 4942)