[�S��]�i173���j ��ژ^�f�ڏ��ɕ\���i1/1)
| |
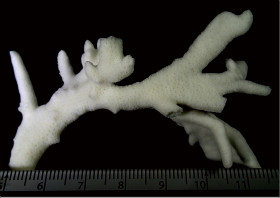
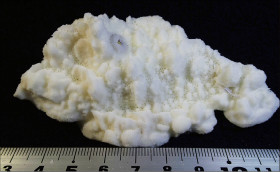
Acropora aculeus (Dana, 1846)
�n���G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[14m)

|

��2���}�̒��a��3�`5mm�B
|
|
�����`�F���}���A�ő��R�����{�[�X��A�ő���Q�́B
��̕��̐F�ʂƓ����F�F�A���F�A���F�A���ΐF��N���[���F�ȂǗl�X�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�ׂ͍��~���`�ŁA�����̂�1�����̏ꍇ�A��߂��̒��a��3�`5mm�ɂȂ�B������3cm�����̂��̂��������A10cm�߂��܂ŐL�т���̂�����B�אڂ����2���}�̒����̊Ԃ̋�����1cm���x�ŁA�Q�̕\�ʂɂقړ��Ԋu�ɕ��ԁB�������A��2���}�̕������ȕ����ł́A���̋����͒Z���Ȃ�B�����̂͂��ꂢ�ȉ~����ŁA�O�a��1.5�`2mm�Ə������A1.5�`3mm���x�ˏo����B���̊O�Ǖ����͔������E���Ɍ����邪�A���ŁB1���u�ǂ̒�����1/3�`1/2R�B���ˌ̂͊O�ǂ����������Ǐ@�`�ŁA�O�a1.1�`1.3mm�قǁA�J�����͉~�`�����`��悷��B1���u�ǂ̒�����1/3�`2/3R�B�����͘]�`�Ԗڏ�ŁA�\�ʂɂ͒P���[�������B����B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[10m �Ȑ[�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Acropora nana �X�Q�~�h���C�V�ƍ�������Ă���A��q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora austera (Dana, 1846)
�R�C�{�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[13m)

|

��2���}�̒��a��1.5cm�O��B
|
|
�����`�F�}���l�X�ȕ��������������}��A�ő������r�u���V��Q�̂������B���[�̐[���Ƃ���ł́A��1���}�����������ɐL�сA���Q�̂̂悤�Ɍ����邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`���F���������A�F��}�̐�[�̐F�ʂ��قȂ����Q�̂�������B
���i�̓����F��3���}�܂Ŕ��B�B��1���}�͒��a1cm�����`��cm�A������cm�`20cm���x�Ɨl�X�B�������̂͊ɂ₩�ɘp�Ȃ���X��������B��2���}�͉~���`�`�~���`�ŁA������a1.5cm�O��A����2�`5cm�̂��̂������B��1���}�Ƒ�2���}�̂Ȃ��p��30�`90���ƕω��ɕx�ށB��3���}�̑����͉~���`�ŁA��ł̒��a�E�����Ƃ���1cm�O��B��3���}�͑�1�E��2���}��ŕs�K���ɔ��B���A���}�Ƃ̂Ȃ��p��60�`90�����炢�B�����̂�䰊O�ǂ������k���ŁA�O�a3�`3.5mm�A1.5�`2mm���x�ˏo����B1���u�ǂ͒���2/3�`3/4R�ł悭���B����B���ˌ͔̂�������Ǐ�ŁA�O�a1.5�`3mm�A�J�����͉~�`�`���p�����~�`�B�����}�̕\�ʂł����ˌ̂̑傫����ˏo�x���͗l�X�ŁA�������Ė��݂������̂���A�����̂̂悤�ɑ傫���A�悭�ˏo�������̂܂ł���B�����͌��łȖԖڏ�ŁA�\�ʂɂ�⑾�߂̍ו���[��(�ꕔ�͒P���[��)�����W����B
�������F�J���I�ȏʎΖʂ̐��[5�`15m�t�߂Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
* ��q�����L�^��

|
|
| |
Acropora cytherea (Dana, 1846)
�n�i�o�`�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[5m)

|

��2���}�̒��a��5�`7mm�B
|
|
�����`�F�������Q�́B��̌��݂�3cm�����B
��̕��̐F�ʂƓ����F��ɒW�ΐF��W���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�ׂ͍��~���`�ŁA�����̂�1�����̏ꍇ�A��ł̒��a5�`7mm�A����1.5�`2.5cm�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm���x�B�����̂͑��E���ł��낭�A�O�a2�`3mm�A1�`2.5mm�قǓˏo����B��̑�2���}�̐�[�ɕ����̒����̂����B������A�Q�̉��ʂɂ���2���}�����B�����肷�邱�Ƃ�����B���ˌ̂͗��`�̍ג����J��������������`�O�ُ�ŁA�O�a��2mm�����ŏ������B�P���u�ǂ͒���2/3R�܂Ŕ��B�B�����́A���ˌ̂̊O���Ř]��A����ȊO�ł͖Ԗڏ�Ŕ��ɂ��낢�B���̕\�ʂɂ́A���Ξ�(�ꕔ�͒P���[��)�����B����B
�������F��2���}�܂Ŕ��B�B��2���}�ׂ͍��~���`�ŁA�����̂�1�����̏ꍇ�A��ł̒��a5�`7mm�A����1.5�`2.5cm�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm���x�B�����̂͑��E���ł��낭�A�O�a2�`3mm�A1�`2.5mm�قǓˏo����B��̑�2���}�̐�[�ɕ����̒����̂����B������A�Q�̉��ʂɂ���2���}�����B�����肷�邱�Ƃ�����B���ˌ̂͗��`�̍ג����J��������������`�O�ُ�ŁA�O�a��2mm�����ŏ������B�P���u�ǂ͒���2/3R�܂Ŕ��B�B�����́A���ˌ̂̊O���Ř]��A����ȊO�ł͖Ԗڏ�Ŕ��ɂ��낢�B���̕\�ʂɂ́A���Ξ�(�ꕔ�͒P���[��)�����B����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{���Acropora hyacinthus �i�����E�~�h���C�V�AA. microclados �}�c�o�~�h���C�V��A. spicifera �N�V�n�_�~�h���C�V�ƍ�������Ă��邽�߁A��q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
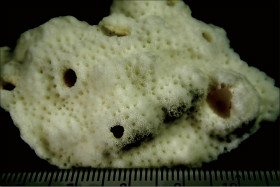
Acropora digitifera (Dana, 1846)
�R���r�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[2m)

|

��2���}�̒��a��1cm�O��B
|
|
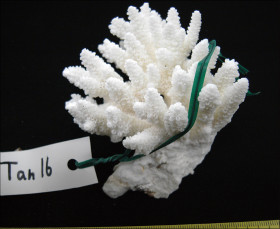
�����`�F�w��A�R�����{�[�X��`�ő��R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F��ɗΐF�⊌�F�A�N���[���F�B�}�̐�[����ˌ̂�䰕NJO���������Ȃ邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�`�~���`�ő傫�����悭�����B�����̂�1�����̑�2���}�́A���a1cm�O��A����2cm�قǁB�אڂ�����2���}�̒����̊Ԃ̋�����1.5cm�O��B�����̂��k���ŊO�a2.5�`3mm�A1�`1.5mm�قǓˏo�B1���u�ǂ͒���1/2�`2/3R�قǁB���ˌ̂͂悭������A�O�a��1.3�`2mm�ŁA�Q�̂��ƂɊO�a��ˏo�x�����悭�����B���ˌ͎̂�Ɍ�����A�Ǐ�`�����Ǐ��A�O�����悭�������̂ŐO�ُ�ɂ�������B�J�����͉~�`�����`�B1���u�ǂ͒���1/2�`2/3R�B�����\�ʂ́A���ˌ̂̊O���ŖԖڏ�`�]��A���ˌ̊Ԃł͖Ԗڏ�ŁA�\�ʂɂ͍ו���[��(�ꕔ�͒P���[��)�����W����B
�������F�g������̋����ʉ�����ʎΖʂ̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F���m�������ȓ�B��q���ł͋H�B
* ��q�����L�^��
|
|
| |
Acropora efflorescens (Dana, 1846) sensu Veron, 2000
�^�C�n�C�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[�s��)

|

��2���}�̒��a��5�`8mm�B
|
|
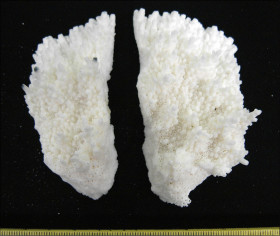
�����`�F�������`��Q�́B��̌���2.5cm�����B�Q�̂����������͂��ꂢ�ȑ�t�^�����A�����ƂƂ��ɂ��̌`�͕���A�Q�̎����������ɒ���o���Ă���B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�̌`��͗l�X�ŁA�����݂̂̂����B�������̂�����B���ˌ̂����B������2���}�͉~���`�~���`�ŁA�傫�����̂Œ��a5�`8mm�A����1cm���x�ƒZ���B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm�����B�����̎}�́A�Q�̂̐����ƂƂ��ɕs���Ă܂��͖��݂��ĕ�����Ȃ��Ȃ邱�Ƃ������B�����̂͑��E���������łŁA�O�a2�`2.5mm�A3�`5mm�قǓˏo�B1���u�ǂ͒���1/2�`2/3R�B���ˌ̂͊Ǐ�`�����Ǐ�ŁA���݂�����̂�����B�O�a1�`2mm�قǂʼn~�`�̊J���������B�����̎��ӂł́A3�`4mm�قǕs�K���ɓˏo���邱�Ƃ�����B1���u�ǂ͒���1/2R�B�����́A���ˌ̂̊O���Ř]��A���ˌ̊Ԃł͖Ԗڏ�ŁA�����̕\�ʂɂ͑��Ξ�(�ꕔ�͒P���[��)�����ԁB
�������F�O�m�ɖʂ����A�܂��͂��Օ����ꂽ��ʈ�̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�`��q���B��q���ł͋H�B
�⑫�FVeron (2000)�͖{���Acropora efflorescens �Ɠ��肵�Ă���BWallace (1999)��Wallace et al. (2012)�́AA. efflorescens ��A. cytherea �n�i�o�`�~�h���C�V�̃V�m�j���Ƃ��Ă��邪�A����͑�2���}�̔z��⒆���̂̌`�Ԃ��قȂ邽�߁A�X�Ȃ镪�ފw�I�������K�v�Ǝv����B�{��̘a���͋��{�C�������Z���^�[(1977)�Ɋ�Â��B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Acropora florida (Dana, 1846)
�T�{�e���~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

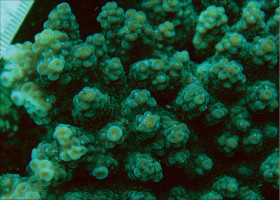
�B�e�F�[���T�L(���V�\�s�匴�A���[8m)
|

��3���}�̒��a��5mm�`1cm�B
|
|
�����`�F�ő���`���r�u���V��A���r�u���V���A���ƌQ�̌`�̕ω����傫���B���Q�̂ł́A��1�E��2���}�Ԃ��Z�����Ăقڔ�ɂȂ������̂�����������B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��3���}�܂Ŕ��B�B��1���}�͒��a1�`3cm�̂��̂��������A����ȏ�̑����ɂȂ邱�Ƃ��������Ȃ��B������10cm�ȏ�ɂȂ邱�Ƃ����ʁB��2���}�͌Q�̎����Ŗ��ĂŁA��ł̒��a1cm�O��A����3�`4cm�܂ŁB��1���}�Ƃ̂Ȃ��p��40�`80���B��3���}�͑����Ⓑ�����悭�����A���a5mm�`1cm�A����1.2�`1.5cm���x�B��1�E��2���}�Ƃ̂Ȃ��p��60���قǁB�אڂ�����3���}�̒����̊Ԃ̋�����1�`1.5cm�B�����̂��k���Ȃ��̂��瑽�E���Ɍ�������̂܂ł���A�O�a�͂قƂ�ǂ�3mm�O��œˏo���Ȃ��B1���u�ǂ͂悭���B���A����2/3R�قǁB���ˌ͖̂����Ǐ�ŊJ�����͉~�`�A�O�a1.5�`2.5mm�A1���u�ǂ̒���1/2�`2/3R�B�����́A���ˌ̂̊O���Ō����Ș]��A����ȊO�ł͖Ԗڏ�ɂȂ镔��������B�\�ʂɂ͒P���[��(�ꕔ�͑��Ξ�)�����B����B
�������F�J���I�ȏʎΖʂ̐��[5�`15m�t�߂Ō����A�ʉ����ʌł��������邱�Ƃ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��̑��Q�̂�Acropora japonica �j�z���~�h���C�V��A. aff.gemmifera �ƍ�������Ă���\��������B��q���Ȗk�ł̐����͍Č������K�v�B
* ��q�����L�^��

|
|
| |
Acropora aff. gemmifera (Brook, 1892)
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[2m)

|

��2���}�̒��a��6mm�`1.2cm�Ƃ���B
|
|
�����`�F�w��`�R�����{�[�X��Q�̂ŁA��Ղ��L�������悤�ɐ�������B�Q�̎����͊�ՂɌŒ������ɒ���o���A�ő��R�����{�[�X��ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɗΐF��Ԋ��F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͎�ɉ~���`�ŁA�Q�̎����ő����ɒ����L�т����͉̂~���`�Ɍ�����B�����Q�̓��ł��}�̑����Ⓑ�����s�����ŁA��͒��a6mm�`1.2cm�A����1�`2.5cm�B�אڂ�����2���}�̒����̊Ԃ̋����͂悭�����A1.2�`1.8cm�قǁB�����̂��k���ŁA�O�a2.5�`3mm�A1mm�قǂ����ˏo���Ȃ��B1���u�ǂ͒���2/3�`3/4R�ŁA䰐S�߂��܂ł悭���B����B���ˌ͉̂~�`�J���ŊǏ�`�����Ǐ�A�O�a1.3�`2mm�̂��̂ƁA1mm�����̂��̂Ƃ�2�^�C�v�����݂���B1���u�ǂ͂��܂蔭�B�����A����1/2R�����B�����\�ʂ́A���ˌ̂̊O���ŖԖڏ�`�]��A���ˌ̊Ԃł͖Ԗڏ�B�����\�ʂ̞��̌`��́A�P���[���A���Ξ���ו���[���ȂǗl�X�B
�������F�g�Q�̉e�����悭��ʉ��A�ʎΖʂ��ʈ�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�Q�̌`����A�{���Acropora japonica �j�z���~�h���C�V�ƍ�������Ă���\��������A�����ł̐����ɂ��Ă͍Č������K�v�B�{��́A��2���}�̌`�����ˌ̂̌`�Ԃ�A. gemmifera �I�����r�~�h���C�V�i�{�K�C�h���f�ڎ�j�Ɩ��炩�ɈقȂ��Ă���B
* ��q�����L�^��

|
|
| |
Acropora glauca (Brook, 1893)
�i�J���r�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

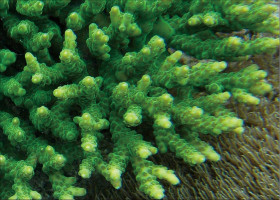
�B�e�F��@��(����q���剖���A���[1m)
|

��2���}�̒��a��5mm�`1cm�B
|
|
�����`�F�w��`�R�����{�[�X��Q�́B����悭���B�����łŁA��Ղɂ�������ƌŒ�����Q�̂������B
��̕��̐F�ʂƓ����F�ΐF�`���ΐF�⊌�F�B��q���ł͕��ʎ�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�`�~���`�ŁA����Q�̓��ł̑����Ⓑ���͕s�����ɂȂ�₷���B�����̂�1�����̑�2���}�́A��ł̒��a5mm�`1cm�A����2�`4cm�قǁB��2���}�̒����⑾�����قړ������Q�̂ł́A�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`1.5cm�Ƃ悭�����B�����̂��k���A���~���`�ŊO�ǂ������A�O�a2.5�`4mm�A1�`3mm�قǓˏo����B1���u�ǂ͒���1/3�`3/4R�B���ˌ͎̂�ɔ�������ۂ������Ǐ�`�@�`�ŁA�O�a1.5�`2.5mm�A�J�����͉~�`�`���`�B�}�̉����ł͖��݂������̂�������B1���u�ǂ͒���2/3R�قǁB�������k���ŁA���ˌ̂̊O���ł͘]��ɂȂ�₷���A���̑��̕����ł͖Ԗڏ�ɂȂ�B�����̕\�ʂɂ͒P���[���`���Ξ����悭���B����B
�������F�O�m�ɖʂ����A�܂��͂��Օ����ꂽ��ʈ�̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��`��q���B��q���ł͕��ʎ�B
�⑫�F�{��́A�Q�̌`�ȊO�̓������ގ�����Acroproa cf.glauca �G���^�N�~�h���C�V�Ƃ̍X�Ȃ镪�ފw�I�������K�v�B�{��̘a���͋��{�C�������Z���^�[ (1977) �Ɋ�Â��B
* ��q�����L�^��
|
|
| |
Acropora cf. glauca (Brook, 1893)
�G���^�N�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���剖���A���[5m)

|
��2���}�̒��a��8mm�`1.2cm�B
|
|
�����`�F���}���Q�́B��̌�����2.5cm�قǁB��1���}�͐��������ɐL�����A�����̊Ԃ������ɗZ�������Q�͔̂�Ɍ�����B��2���}�͒Z���A���̊Ԋu���悭�����̂ŁA�Q�̂͂��ꂢ�ȉ~��^�Ɍ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B��q���ł͕��ʎ�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�`�~���`�ŁA�����̂�1�����̏ꍇ�A��ł̒��a8mm�`1.2cm�A����2.5cm�����B�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`2cm�ł悭�����B�����̂��k���A���~���`�ŊO�ǂ������A�O�a2.5�`4mm�A1�`2mm�قǓˏo����B1���u�ǂ͂悭���B���A����2/3R�قǁB���ˌ͔̂�������ۂ������Ǐ�`�@�`�ŁA�O�a1.5�`2.5mm�A�J�����͉~�`�`���`�B�}�̐�[�߂��ł́A�s�K���ɓˏo�����Ǐ�̂�������B1���u�ǂ̒���1/3�`1/2R�O��B�������k���ŁA���ˌ̂̊O���ł͘]��ɂȂ�₷���B���̑��̕����ł͖Ԗڏ�ɂȂ�A�\�ʂɂ͒P���[�������W����B
�������F�O�m�ɖʂ����A�܂��͂��Օ����ꂽ��ʈ�̐��[3�`15m�Ō�����B
�����ł̕��z�F��t���َR�E���茧�Δn�`��q���B��q���ł͕��ʎ�B
�⑫�F�{��́A�Q�̌`�������������ގ�����Acropora glauca �i�J���r�~�h���C�V�Ƃ̍X�Ȃ镪�ފw�I�������K�v�B�{��̘a���͋��{�C�������Z���^�[ (1977) �Ɋ�Â��B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora horrida (Dana, 1846)
���Z�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[14m)

|

��2���}�̒��a��7�`9mm�B
|
|
�����`�F���}��A�ő���܂��͐��r�u���V��ɂȂ�B���[�̐[���Ƃ���ł͑�1���}�����������ɐL�тāA���r�u���V���Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F�D�F���������ΐF�`���F�ŁA�Q�̂ɂ���ĐF�ʂ̔Z�����قȂ�B���Ԃ��G���L���Ă��邱�Ƃ������B
���i�̓����F��3���}�����B�B�Q�̎����ł̑�1���}�Ƒ�2���}�̒��a�͂��ꂼ��1�`1.2cm��7�`9mm�B��2���}�̒����͕s�����ŁA���̑�����5cm�����B��1���}�Ƒ�2���}�̂Ȃ��p�͑傫���A60�`80���ɂȂ�B��3���}�͑�1�E��2���}�̗����Ɍ����A��1���}��ł̔z�u�͕s�K���ŁA�������s���������A5mm�`1.2cm���炢�̂��̂������B��1�E��2���}�Ƒ�3���}�̂Ȃ��p�͂Ƃ���40�`90���B��2�E��3���}�͊ɂ₩�ɘp�Ȃ��Ă�����̂������B�����̂͑��E���ŁA1���u�ǂ�2/3R�܂Ŕ��B�B��2���}�ł̒����̂̊O�a��2�`2.5mm�A�ˏo���Ȃ����̂���4mm�߂��ˏo������̂܂ŗl�X�B���ˌ̂͊O�a2.5mm���x�ŁA�ˏo�x��������A�Ԋu�͕s�����B��3���}�ł͕��ˌ̂̐������Ȃ��A���܂�ˏo���Ȃ��B���ˌ̂͊Ǐ�ʼn~�`�J���̂��̂��������A�}�̐�[�߂��ł͕@�`�ŗ��`�̊J�����������̂�����B1���u�ǂ͒���1/2R�O��B�����͖Ԗڏ���k���A�\�ʂɂ͒P���[�������B����B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�[��ł͑�Q�����`�����邱�Ƃ������B
�����ł̕��z�F��q���ȓ�B��q���ł͋ɂ߂ċH�B
* ��q�����L�^��

|
|
| |
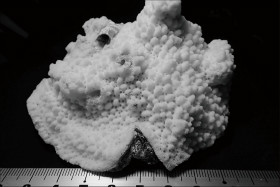
Acropora humilis (Dana, 1846)
�c�c���r�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�����@�O(���V�\�s��Óc�A10m)

|

��2���}�̒��a��1.5�`2cm�B
|
|
�����`�F�w��`�R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͑����~���`���~���`�A��͎O�p�`�l�p�`�ɂȂ��Ă��邱�Ƃ������B�����̂�1�����̑�2���}�͒��a1.5�`2cm�A����2�`2.5cm�B�אڂ�����2���}�̒����̊Ԃ̋�����2cm�����B�����̂��k���ŁA�O�a��4�`5mm�Ƒ傫���A�ˏo���Ȃ��B1���u�ǂ��悭���B���A����3/4R�قǁB���ˌ̂͑傫���A�O�a1.7�`2mm�B��ɊǏ�ŁA������̂��̂����邪�A�ˏo�̒��x������͂悭�����B�J�����͗��`����~�`�B1���u�ǂ̒����͗l�X�ŁA1/2R�����`3/4R�B�����\�ʂɂ͑��Ξ��܂��͍ו���[�������W���A���ˌ̂̊O���ł͂���炪�z�Ę]���`�����邱�Ƃ�����B
�������F�g������̋����ʉ�����ʎΖʂ̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
* ��q�����L�^��

|
|
| |
Acropora hyacinthus (Dana, 1846)
�i�����E�~�h���C�V�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[2m)

|

��2���}�̒��a��5�`7mm�B
|
|
�����`�F���Q�́B��̍�����3cm�قǁB
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ő傫�����悭�����A�����̂�1�����̏ꍇ�A��Œ��a5�`7mm�A����1.5cm�����̂��̂��قƂ�ǁB�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm�����A��[�ɒ����̂���������}�������ƁA�Ԋu�͂�苷��������B�����̂͑��E���A�O�a�͏�����1.5mm�قǂŁA�ˏo���Ȃ��B1���u�ǂ̒�����1/2�`2/3R�B���ˌ̂͊O�a1�`1.5mm�̌�����`�O�ُ�ŁA�J�����͂قڐ����ʼn~�`�Ɍ�����B1���u�ǂ̔��B�������A����1/2R�܂ŁB�����͕��ˌ̂̊O���ł͘]��A���ˌ̊Ԃł͖Ԗڏ�B�\�ʂɂ͒P���[���`���Ξ������B����B
�������F�g������̋����ʉ�����ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Acropora cytherea �n�i�o�`�~�h���C�V��A. spicifera �N�V�n�_�~�h���C�V�ƍ�������Ă��邽�߁A��q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B�{��̐V�̘a���́A�{�킪��q���ȓ�̃T���S�ʈ���\����~�h���C�V���̈��ł��邱�ƂɗR������B
* ��q�����L�^��

|
|
| |
Acropora intermedia (Brook, 1891)
�g�Q�X�M�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[4m)

|

��2���}�̒��a��1.5�`2cm�B
|
|
�����`�F���}��Q��
��̕��̐F�ʂƓ����F���F�A�N���[���F��F�B
���i�̓����F��2���}�܂Ŕ��B�B��1���}��20cm�ȏ�L�сA�}�̉����ʼn~���`�A��[10cm�قǂ͐�[�Ɍ������ĉ~���`�ɂȂ�B��[����10cm�t�߂̒��a�͖�1.5�`2cm�B��1���}��20 cm�ȏ�ɐL�т����̂̒��a�́A����ȏ�̑����ɂȂ邱�Ƃ�����B��2���}�͉~���`�ŁA��1���}�̐�[����10cm�قǂ̊Ԃł悭���B����B������10cm�����A��ł̒��a��8mm�`1.5cm�قǁB��1���}�Ƒ�2���}�̂Ȃ��p��50�`80���B�����̂͑��E���ŁA�O�a3�`4mm�A2�`3mm�قǓˏo����B1���u�ǂ̒�����1/3�`2/3R�B���ˌ̂ɂ͎���2�^�C�v������B(1)�O�a1�`2.5mm�Ŏߏ������1.5�`5mm�قǓˏo�����Ǐ�A������܂��͕@�`�ɂȂ������́B(2) �O�a1�`1.5mm�Ŗ��݂܂��͖����Ǐ�ɂȂ������́B��Ɍ�҂͑O�҂̊�t�߂ɔ��B����B�O�҂͉~�`�����`�A��҂͉~�`�̊J���������B���ˌ̂�1���u�ǂ͒���1/2�`2/3R�B�����́A���ˌ̂̊O���Ř]��A���ˌ̊Ԃł͖Ԗڏ�ŁA�Ƃ��ɕ\�ʂɂ͒P���[�������B����B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋ɂ߂ċH�B
�⑫�F�{���Acropora muricata �X�M�m�L�~�h���C�V�ȂǑ��̎��}��~�h���C�V�ނƍ�������Ă��邽�߁A��q���ȓ�ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Acropora japonica Veron, 2000
�j�z���~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[3m)

|

��2���}�̒��a��5mm�`1.5cm�ŕs�����B
|
|
�����`�F�w��`�R�����{�[�X��Q�̂ŁA��Ղ��悤�ɐ�������B���p�␅�[10m�Ȑ[�̔g�Q�̎ア�����ł́A�ő��R�����{�[�X��Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F���ΐF��ΐF�̂��̂������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ŁA�Q�̎����ő����ɒ����L�т����͉̂~���`�Ɍ�����B�����Ⓑ���͕s�����ŁA�}��͒��a5mm�`1.5cm�A����5mm�`2.5cm�B�אڂ����2���}�̒����̊Ԃ̋������s�����ŁA1cm�����̂Ƃ����3cm�߂�����Ă���Ƃ��낪���݁B�����̂��k���ŁA�ʏ�͂��܂�ˏo���Ȃ����A�����H��1.5mm�قǓˏo���邱�Ƃ�����B�O�a��2�`3mm�̂��̂��������A�������ĕ��ˌ̂Ƌ�ʂ����Ȃ����̂�����B1���u�ǂ͂悭���B���A����2/3�`3/4R�B���ˌ͔̂�����������Ǐ�ŁA�J�����͉~�`�ŏ������A�O�a��1.5�`2mm�̂��̂ƁA1.2mm�����̂��̂�2�^�C�v��������B�����2�^�C�v�̔z��͕s�K���Ȃ��߁A�}�\�ʂ͂ł��ڂ����Č�����B1���u�ǂ̒�����1/2R�قǁB�����͑e���Ԗڏ�ŁA�\�ʂɂ͒P���[���⑤�Ξ������ԁB
�������F�g�Q�̉e�����悭���ʈ�̐��[5m�Ȑ�ł悭������B
�����ł̕��z�F�a�̎R�����{�E���茧�Δn�`��q���B��q���ł͕��ʎ�B
�⑫�F�{��̃^�C�v�Y�n�͋{�茧�����s���Y�B�a���͖쑺�E�ڍ�(2005)�Ɋ�Â��B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FA. striata�Ƃ��ċL�^�����\������

|
|
| |
Acropora cf. latistella (Brook, 1892)
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�����@�O(���V�\�s���Y�A���[15m)

|

��2���}�̒��a��4�`5mm�B
|
|
�����`�F���}���`�ő��R�����{�[�X��Q�̂ō�����5cm�ɖ����Ȃ��B��1���}�Ԃ��Z�����Ĕ�Q�̂ɂȂ邱�Ƃ�����B�Q�̎����ł͑�2���}�̔��B�������A��1���}�������ɒ���o�����悤�Ɍ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Q�̂ɂ���ĐF�ʂ̔Z�����قȂ�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ŁA�����̂�1�����̏ꍇ�A�傫�����̂͒��a4�`5mm�A����2.5�`4cm�ɂȂ�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm�����B�����̂͑��E���������łŁA�O�a����2.2mm�A�����2mm���x�ˏo����B1���u�ǂ́A䰐S�߂�(3/4R)�܂Ŕ��B���邱�Ƃ������B���ˌ̂́A�O�a2mm�����ŏ������A�~�`�J���������A1���u�ǂ�2/3R�܂Ŕ��B����B�O�`�͎�ɖ����Ǐ�ŁA�}�̏���ł͐O�ُ�ɋ߂��B�����̌`��́A���ˌ̂̊O�����A�אڂ�����ˌ̊Ԃ��]��ŁA�P���[���ŕ�����B
�������F�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō����A�J���I�Șp�ł͂��ł�������B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�{���Acropora latistella �L�N�n�i�K�T�~�h���C�V�i�{�K�C�h���f�ڎ�j�Ɏ��邪�A��2���}�̒��a�⋤���̌`�قȂ邽�߁A�X�Ȃ镪�ފw�I�������K�v�B�܂��AA. cytherea �n�i�o�`�~�h���C�V��A. microclados �}�c�o�~�h���C�V�ƍ�������Ă��邽�߁A��q���Ȗk�ł̐����̍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Acropora microclados (Ehrenberg, 1834)
�}�c�o�~�h���C�V�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(���V�\�s��Óc�A���[5m)

|

��2���}�̒��a��5�`7mm�B
|
|
�����`�F���}���`�ő��R�����{�[�X��Q�́B��̍�����5cm���x�B
��̕��̐F�ʂƓ����F��ɖ��邢�ΐF�⊌�F�B���Ԃ��G����L���Ă��邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͎�ɍׂ��~���`�ŁA�����̕��ˌ̂��悭����o�����}�͂��~���`�ɂ݂���B�����̂�1�����̑�2���}�̒��a�́A���5�`7mm�A��[�߂���3�`5mm�A������1.5�`2cm�ɂȂ�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`1.5cm�B��̑�2���}�̐�[�ɁA�����̂悭�ˏo���������̂ƕ��ˌ̂����B���邱�Ƃ������A���̏ꍇ�͎}�̒��a�͂�葾���A�}�̊Ԋu�͋����Ȃ�B��2���}�͌Q�̂̉��ʂɂ��悭���B���邱�Ƃ�����B�����̂͑��E���A�O�a2�`3mm�ŁA���܂�ˏo���Ȃ����̂���5mm�ȏ�ˏo������̂܂ŗl�X�B1���u�ǂ�2/3R�܂Ŕ��B����B���ˌ͉̂~�`�`���`�J���̔����O�ُ��@�`�ŁA�ߏ�Ɍ������Ă悭����o���B�O�a2mm�����A1���u�ǂ̔��B�͈����B�����́A���ˌ̂̊O���Ř]��A���ˌ̊ԂŖԖڏ�A���̕\�ʂɂ͑��Ξ������ԁB
�������F�J���I�ȏʎΖʂ̐��[5�`15m�t�߂Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B
�⑫�F�{���Acropora cytherea �n�i�o�`�~�h���C�V��A. latistella �L�N�n�i�K�T�~�h���C�V�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B�{��̐V�̘a���́A��[���ׂ��}�����ꂵ����2���}�����t�Ɏ��Ă��邱�ƂɗR������B
* ��q�����L�^��
|
|
| |
Acropora muricata (Linnaeus, 1758)
�X�M�m�L�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���剖���A1m)

|

��2���}�̒��a��1.2cm���x�B
|
|
�����`�F���}��`�ő���Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�}�S�̂܂��͎}�̐�[���݂̂��F�����������̂�����B
���i�̓����F��2���}�܂Ŕ��B�B��1���}��20cm�ȏ�L�сA�����ł͉~���`�A��[10cm�قǂ͉~���`�ɂȂ�B��[����10cm�t�߂ł̒��a��1.2cm���x�B��2���}�͉~���`�ŁA��1���}�̐�[����10cm�قǂ̊Ԃł悭���B���A������5cm�����A��ł̒��a��1cm�����̂��̂������B��1���}�Ƒ�2���}�̂Ȃ��p�͒ʏ�50�`80�����炢�ŁA���܂�90��������̂�����B�����̂͑��E���ł͂Ȃ����A���낭�ĂԂ�₷���B�O�a2.5�`3.5mm�ł��܂�ˏo����(2mm����)�A1���u�ǂ�1/2�`3/4R�܂Ŕ��B�B���ˌ̂ɂ�2�^�C�v����A��͉~�`�J���̊Ǐ�`�����Ǐ�A�O�a1�`2.5mm�B������͉~�`�J���̖��݁`�����Ǐ�A�O�a0.5�`1.5mm�B��҂͑O�҂̊�߂��ł悭���B����B1���u�ǂ̒����͗��^�C�v�Ƃ�1/2�`2/3R�قǁB�����́A���ˌ̂̊O���Ř]��A����ȊO�̂Ƃ���ł͖Ԗڏ�ɂȂ�A�\�ʂɂ͒P���[���⑤�Ξ���������B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʈ�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Acropora intermedia �g�Q�X�M�~�h���C�V�ȂǑ��̎��}��~�h���C�V�ނƍ�������Ă��邽�߁A��q���ȓ�ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��

|
|
| |
Acropora nana (Studer, 1878)
�X�Q�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[5m)

|
��2���}�̒��a��4�`6mm�B
|
|
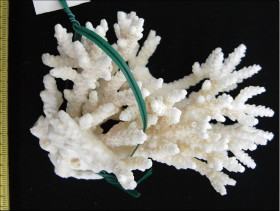
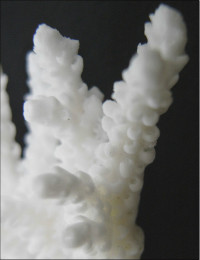
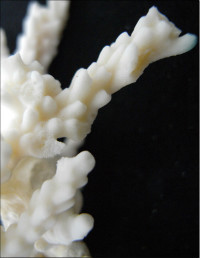
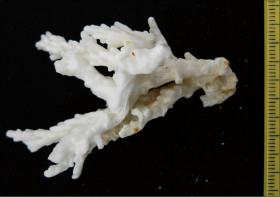
�����`�F�@�ׂȊO�ς̎��}���A�ő��R�����{�[�X��A�ő���Q�́B
��̕��̐F�ʂƓ����F�ΐF�A���F�A�N���[���F�A�F�⎇�F�ȂǗl�X�B�}�̐�[�̓s���N�F�Ȃǂ̒W�F�ɂȂ邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�ׂ͍��~���`�ŁA�����̂�1�����̏ꍇ�A���a4�`6mm�A����3cm�قǂ̂��̂�������10cm�߂��܂ŐL�т���̂�����B�אڂ����2���}�̒����̊Ԃ̋�����1.2�`1.5cm���x�ŁA�قړ��Ԋu�ɕ��ԁB�������A��2���}�̕������ȕ����ł́A���̋����͂��Z���B�����̂��k���ŊO�ǂ͌����A��[�����ۂ܂����~���`�B�O�a1.5�`2mm�A1.5�`2.5mm�قǓˏo�B1���u�ǂ͒���2/3�`4/3R�B���ˌ̂́A�~�`�J���̖����Ǐ�ŁA�ۂ����������̂�����B�O�a1.5�`2mm�B1���u�ǂ͒���1/2�`2/3R�B�����͌��łȖԖڏ�ŁA�\�ʂɂ͍ו���[��(�ꕔ�͒P���[��)�����W����B
�������F���ʂ��̂悢�ʉ�����ʎΖʂ̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B
�⑫�F�{��́A���l�̌Q�̌`�ɂȂ�Acropora aculeus �n���G�_�~�h���C�V��A. valida �z�\�G�_�~�h���C�V�ƍ�������Ă���B�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Acropora nasuta (Dana, 1846)
�n�i�K�T�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A2m)

|

��2���}�̒��a��8mm�`1cm�B
|
|
�����`�F������5cm�قǂ̃R�����{�[�X��`�ő��R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͐�[�̂ق�����⑾���~���`�ŁA�����̂�1�����̏ꍇ�A��Œ��a8mm�`1cm�A����3cm�قǁB�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`1.5cm�B�����̂��k���ŁA�O�a2�`2.5mm�A1mm�قǂ����ˏo���Ȃ��B1���u�ǂ͒���1/2�`2/3R�B���ˌ̂͊O�a1.5�`2mm�B�}�̏���ł͗��`�J���̕@�`�ł悭����o�����̂������A�����ł͉~�`�J���Ŗ����Ǐ�̌̂������B���̂��߁A��2���}��^�ォ�猩��ƁA�����̒����̕��ˌ̂��悭�ڗ��B�������k���ŁA���ˌ̂̊O�����]��Ɍ����邪�A�S�̓I�ɂ͖Ԗڏ�ŁA�\�ʂ𑽐��̒P���[���������Ă���B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
*** ����܂Ŏ�q���Ŋm�F����Ă��Ȃ���

|
|
| |
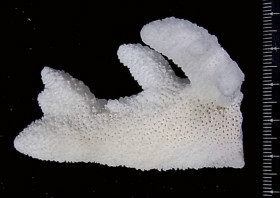
Acropora palmerae Wells, 1954
�}�c�J�T�~�h���C�V�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�쑺�b��(����q���剖���A���[2m)
|

��2���}�̒��a��1�`3cm�Ƃ���B
|
|
�����`�F�핢��`�w��Q�́B�����̕s�����ȑ�2���}���s�K���ɔ��B���邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B���邪�A�����ꍇ������B�܂��A��2���}��[�ɒ����̂��`������Ă��Ȃ����̂�������B�悭���B������2���}�͉~���`�`�~���`�ŁA���a1�`3cm�A����10cm�����B�����̂��k���ŊO�a3mm�O��A�ˏo���Ȃ��B1���u�ǂ͒���1/2�`3/4R�ŁA���Q�̓��ł��̂ɂ���ėl�X�B���ˌ̂ɂ́A�O�a2mm�O��Ō����J���̊Ǐ�`�悭�J�����O�ُ�̂��̂ƁA1mm�O��̖����Ǐ�`���݂������̂�2�^�C�v�������A����炪�Q�̕\�ʂɖ��W���ĕ��z����B���^�C�v�Ƃ�1���u�ǂ̔��B�������A�����Ă�1/2R�܂ŁB�����́A���ˌ̂̑��ʂ��]��ɂȂ邱�Ƃ����邪�A�啔���͖Ԗڏ�ŁA���̔��B�������\�ʂ͕����Ɍ�����B
�������F�g������̔��ɋ����ʉ�����ʎΖʂ̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F��q���ȓ�B
�⑫�F�{���Acropora aborotanoides �g�Q�}�c�~�h���C�V��A. robusta ���X���~�h���C�V�i����Ƃ��{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�B�{��̐V�̘a���́A��������Č�������ˌ̂ɂ���āA�Q�̕\�ʂ����ڂ�����̕\�ʂɎ��邱�ƂɗR������B
* ��q�����L�^��
|
|
| |
Acropora papillare Latypov, 1992
�^�P�m�R�~�h���C�V�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F���r�a�K(����q���n���̊≮�A���[1m)
|
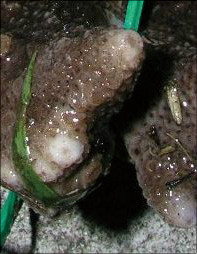
��2���}�̒��a��2cm�O��B
|
|
�����`�F��Ɏw��Q�́B�Q�̎����̎}�������ɐL�������R�����{�[�X��Q�̂�������B
��̕��̐F�ʂƓ����F��Ɋ��F�`�W���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�`�~���`�B�܂�Ɏ}�̑����A������Ԋu���悭�����Q�̂����邪�A�ʏ�͂���炪�s�����ȌQ�̂̂ق��������B�}��Œ��a2cm�O��A����2.5�`3.5cm�B�אڂ�����2���}�̒����̊Ԃ̋����́A�}�����W���Ă���Ƃ����2�`3cm�قǁB�����̂��k���A�O�a3mm�O��œˏo���Ȃ��B�P���u�ǂ͒���1/3�`3/4R�B���ˌ̂͊O�a1.5�`1.8mm�ő傫�����悭�����A��2���}�̑��ʂł͖��W���ĕ��z����B���̌`��͎�ɐO�ُ�ŁA���O�͎}�̏㕔�ł͔����A�����ł͂�������Ċۂ݂�ттČ�����B�P���u�ǂ̔��B�͈����A����1/4�`1/3R�B�����́A���ˌ̂̊O���Ř]��A���̑��̕����ł͖Ԗڏ�ŁA�\�ʂɒP���[��(�ꕔ�͑��Ξ�)�����W����B
�������F�g������̔��ɋ����ʉ��`�ʎΖʂ̐�(���ɐ��[5m�Ȑ�)�Ō�����B
�����ł̕��z�F��q���ȓ�B
�⑫�F�{���Acropora aspera �q���}�c�~�h���C�V�AA. monticulosa �T���J�N�~�h���C�V��A. robusta ���X���~�h���C�V�i������3��͑S�Ė{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B�{��̐V�̘a���́A�}�̊O�ς��^�P�m�R�Ɏ��邱�ƂɗR������B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FAcropora sp. Japan 2�Ƃ��ċL�^�����\������
|
|
| |
Acropora pruinosa (Brook, 1892)
�q���G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F���n�m�V(�É������ɓ����c�q�A���[5m)

|
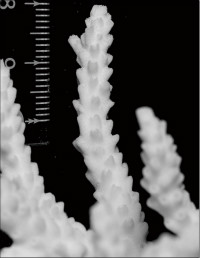
��2���}�̒��a��6�`8mm�B
|
|
�����`�F�ő���`���}��Q�̂ŁA�Q�̂̍�����20cm���邱�Ƃ�����B�J���I�Șp�̊�Տ�ł́A������10cm�ɖ����Ȃ��ő��R�����{�[�X��Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B���ΐF�Ȃǂ̖��邢�F�ʂ̌Q�̂�������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͒�����5cm�����ŁA�}�̉����ł͉~���`�A��[1cm�t�߂���ׂ͍��Ȃ��ĉ~���`�ɂȂ�B�܂����}��Q�̂̏ꍇ�A�����̑�2���}�͊����1cm���炢�̂Ƃ���Řp�Ȃ��Ă��邱�Ƃ������B�����̂�1�����̑�2���}�ł́A��Œ��a6�`8mm�B���}��Q�̂̏ꍇ�A��1���}��10cm�ȏ�L�сA��[����8�`10cm�t�߂Œ��a8mm�`1cm�ɂȂ�B��[����5cm�قǂ̌`��͑�2���}�Ƃقړ����B��1���}�Ƒ�2���}�Ƃ̂Ȃ��p��50�`70���B�����̂͑��E���ŁA�O�a2�`3mm�A�ˏo���x��2mm�����B1���u�ǂ��悭���B���A����2/3�`1R�B���ˌ̂͒����Ǐ�ŁA�O�a2mm�O��B�J�����͂��ꂢ�ȉ~�`�ŁA1���u�ǂ͒���2/3�`1R�B�����͖Ԗڏ�ŁA���̕\�ʂɒP���[���⑤�Ξ������W����B���ˌ̂̑��ʂł́A�����̞������ɕ���Ř]��ɂȂ邱�Ƃ������B
�������F���[10m�Ȑ�̎Օ��I�Șp�̍��I���A�J���I�Șp�̐��[5�`10m�̊�Տ�Ȃǂő�Q�����`�����₷���B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�A���������m�ȕ��z����͕s���B��q���ł͋L�^����Ă��Ȃ��B
�⑫�F�{��͂���܂�Acropora tumida �G�_�~�h���C�V�ƍ�������Ă����B�{��̃^�C�v�Y�n�͒��茧�Δn�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora secale (Studer, 1878)
�g�Q�z�\�G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�[���T�L(���V�\�s�匴�A���[9m)

|

��2���}�̒��a��8mm�`1cm�B
|
|
�����`�F�R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F��Ɋ��F�ŁA�}�̐�[�����F�ɂȂ邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͐�[����⑾���~���`�B�����̂�1�����̑�2���}�ł́A��Œ��a8mm�`1cm�A����2.5cm�قǁB�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`1.5cm�B�����̂��k���ŊO�ǂ������A��[�����ۂ܂����~���`�A�O�a3mm���x�A1�`2.5mm�قǓˏo�B1���u�ǂ͒���1/2�`2/3R�B���ˌ̂ɂ�2�^�C�v����A��͊O�a1.8�`2.5mm�̔�������@��`�Ǐ�ŁA���`�`�~�`�J���B������͊O�a1�`1.2mm�̖��݁`�����Ǐ�ʼn~�`�J���B���^�C�v�Ƃ�1���u�ǂ͒���1/3�`2/3R�B�����͌��łȖԖڏ�ŁA�\�ʂɂ͍ו���[������������B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
* ��q�����L�^��

|
|
| |
Acropora cf. selago (Studer, 1878)
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���剖���A���[1m)

|

��2���}�͒��a��7�`9mm�B
|
|
�����`�F����5cm�����̃R�����{�[�X��`�ő��R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F��Ɋ��F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ő傫�����悭�����A�����̂�1�����̑�2���}�ł́A���a7�`9mm�A����2�`3cm�B�אڂ�����2���}�̒����̊Ԃ̋�����1.5cm�O��B�����̂��k���ŊO�a2�`2.5mm�A���܂�ˏo���Ȃ��B1���u�ǂ͒���1/2�`2/3R�قǁB���ˌ̂͌�����`�O�ُ�A�J�����͉~�`�`���`�ŁA�O�a1�`2mm�A�悭����o���̂ƂقƂ�ǒ���o���Ȃ��̂�2�^�C�v������B1���u�ǂ͒���1/3�`1/2R�B�����\�ʂ́A���ˌ̂̊O���ł͘]��A���ˌ̊Ԃł͖Ԗڏ�ŁA���̕\�ʂɂ͒P���[��������B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�ʉ�����ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͐����EVeron (1995) ��Acropora sp. Japan 2 �Ƃ��Čf�ڂ���Ă���킾�Ǝv����B�{��́AAcropora selago �^�`�n�i�K�T�~�h���C�V�i�{�K�C�h���f�ڎ�j�̂ق��AA. digitifera �R���r�~�h���C�V��A. millepora �n�C�}�c�~�h���C�V�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���\��������A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora solitaryensis Veron and Wallace, 1984
�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���剖���A���[3m)

|

��2���}�̒��a��1cm�܂ŁB
|
|
�����`�F����5cm������}���`�ő��R�����{�[�X��Q�́B�����ɐL�т���1���}�Ԃ������I�ɗZ�����Ĕ�Q�̂Ɍ����邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�́A�Q�̂ɂ���ĉ~���`�ɂȂ���̂Ɖ~���`�ɂȂ���̂�����A��[������ɕ��邱�Ƃ�����B�}��[�����Ă��Ȃ���2���}�̒�����4cm���x�B�����̂�1�����̑�2���}�ł́A��ł̒��a�́A�~���`�̂��̂�7�`9mm�A�~���`�̂��̂���8mm�`1cm�ł�⑾�߁B�אڂ�����2���}�̒����̊Ԃ̋����́A��[�̕����Ȃ��A�������悭�����Ă���Q�̂�1.5�`2cm���x�B�����̂��k���ŁA�O�a2�`2.5mm�A1�`1.5mm���x�ˏo�B�ꎟ�u�ǂ͒���1/3�`1/2R�B���ˌ̂͊O�a1.5�`2.5mm�Ƒ傫�߂ŁA�~�`�J���̕@�`�܂��͖����Ǐ�A��1�u�ǂ͒���1/2R�قǁB�����́A���ˌ̂̊O���Ř]��A���̑��̕����ł͖Ԗڏ�ŁA�\�ʂɑ��Ξ�(�ꕔ�͒P���[��)���悭���B����B
�������F�O�m�ɖʂ����A�܂��͂��Օ����ꂽ�ʎΖʂ��ʈ�̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F��t���َR�E���茧�Δn�`����{���k���B��q���ł͕��ʎ�B
�⑫�F�{��ɂ́A��2���}�ׂ̍����̂Ƒ������̂�2�^�C�v���܂܂�邱�Ƃ���A�X�Ȃ镪�ފw�I�������K�v�Ǝv����B�܂��{��́A��q���Ȗk�ł�Acropora japonica �j�z���~�h���C�V��A. cf.glauca �G���^�N�~�h���C�V�ƁA��q���ȓ�ł�A. cerealis ���M�m�z�~�h���C�V�i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���\��������B�{��̘a���͔���(1970)�Ɋ�Â��B
* ��q�����L�^��
|
|
| |
Acropora spicifera (Dana, 1846)
�N�V�n�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���剖���A���[4m)

|

��2���}�̒��a��4�`5mm�B
|
|
�����`�F�������`���}���Q�̂ŁA��̌��݂�2cm�����B�אڂ����1���}���m���Z�����邱�Ƃ����邪�A���S�Ȕ�ɂ͂Ȃ�Ȃ��B
��̕��̐F�ʂƓ����F�ΐF�`���F�B���Ԃ��G����L���Ă��邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�ׂ͍��Z���~���`�`�~���`�ŁA�����̂�1�����̏ꍇ�A�傫�����̂Œ��a4�`5mm�A����7mm�`1.2cm�قǂɂȂ�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm�����B�����̂͑��E���ŁA�O�a��1.5�`2mm�B�Q�̎����̂��̂������Ώ���ɓˏo���邱�Ƃ͂Ȃ��B1���u�ǂ͒���1/2�`2/3R�قǁB���ˌ̂͊O�a2mm�����ŁA1���u�ǂ̔��B�������B�`��͎�Ɍ�����`�O�ُ�A�J�����̉��͂قڐ��������A���O���悭����������ˌ̂͂��ߏ�ɒ���o�����悤�Ɍ�����B�����́A���ˌ̂̊O���Ř]��A���ˌ̊Ԃł͖Ԗڏ�ŁA�\�ʂɂ͑��Ξ������ԁB
�������F���ʂ��̂悢�ʒr�E�ʌ�A�ʉ��`�ʎΖʁA��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���ȓ�B
�⑫�F�{���Acropora hyacinthus �i�����E�~�h���C�V�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
*** ����܂Ŏ�q���Ŋm�F����Ă��Ȃ���
|
|
| |
Acropora tanegashimensis Veron, 1990
�^�l�K�V�}�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[2m)

|

��2���}�̒��a��6�`8mm�B
|
|
�����`�F>������1.5cm���x�̔������Q�̂ŁA��1���}�Ԃ������I�ɗZ�����Ĕ�Ɍ�����Ƃ��낪����B
��̕��̐F�ʂƓ����F�Â��ΐF�`���F�ŁA���Ԃ��G���L���Ă��邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ŁA�����̂�1�����̑�2���}�̏ꍇ�A�傫�����̂Œ��a6�`8mm�A����5�`8mm�B�אڂ�����2���}�̒����̊Ԃ̋�����1.2cm�����B�����̂͑��E���ŁA�O�a1.5mm�O��Ə������ˏo���Ȃ����߁A���ˌ̂Ƌ�ʂ����ɂ����B�����̂�1���u�ǂ͒���2/3R���x�B���ˌ̂́A�O�a�������̂Ƃقړ��������傫�߂�1.5�`2mm�B�O�`�͌�����`�O�ُ�ŁA���O�������ɂ悭����o���B���ˌ̂́A�����u�ǂ�䰐S�߂�(3/4R)�܂Ŕ��B���邪�A����ȊO��1���u�ǂ̔��B�͂悭�Ȃ�(1/4R�`1/3R)�B�����́A���ˌ̂̊O���Ř]��A���ˌ̊Ԃł͖Ԗڏ�ŁA�\�ʂɂ͒P���[���⑤�Ξ������ԁB
�������F�J���I�A�܂��͂��Օ����ꂽ��ʈ�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�`��q���B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͎�q�����V�\�s�Z�g�B�{���Acropora hyacinthus �i�����E�~�h���C�V��A. spicifera �N�V�n�_�~�h���C�V�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora tenuis (Dana, 1846)
�E�X�G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(���V�\�s�Y�c�p�A���[5m)

|

��2���}�̒��a��8mm�O��B
|
|
�����`�F���}���`�R�����{�[�X�ő���Q�̂ŁA��̍�����5cm�قǂɂȂ�B
��̕��̐F�ʂƓ����F�ΐF�`���F�ŁA���ˌ̂̉��O���W�F�ʼn�����邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ŁA�����̂�1�����̑�2���}�̏ꍇ�A��Œ��a8mm�O��A����3�`4cm�̂��̂������B�אڂ�����2���}�̒����̊Ԃ̋�����1.2�`1.5cm�قǂł悭�����B�����̂͑��E���ŁA�O�a2.5�`3mm�A1�`3mm�قǓˏo����B1���u�ǂ͒���1/3R�قǁB���ˌ͎̂��`�`�O�ُ�ʼn������悭����o���A�J�����͉~�`�Ɍ�����B���̂��ߑ�2���}��^�ォ�猩��Ƃ��ꂢ�ȃ��[�b�^��Ɍ�����B�O�a1.5�`2.5mm�A1���u�ǂ͒���1/2�`2/3R�B�����́A���ˌ̂̊O���ł��ꂢ�Ș]��A���ˌ̊Ԃł͖Ԗڏ�ɂȂ�B�\�ʂɂ͒P���[�������B����B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋ɂ߂ċH�B
** ����̒����ł͊m�F�ł��Ȃ������������EVeron(1995)�ł͎�q���Ŋm�F����Ă����
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acropora tumida (Verrill, 1866)
�G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F�����@�O(���m���匎�������A���[3m)

|

��2���}�̒��a��8mm�`1cm�B
|
|
�����`�F�w��܂��̓R�����{�[�X��Q�́B��1���}������ɐL�тĎő��R�����{�[�X��`�ő���Q�̂ɂȂ���̂�����B
��̕��̐F�ʂƓ����F��ɒW���F�`���ΐF�ŁA���F��ΐF�̂��̂�������B�����̂̓N���[���F�ɂȂ邱�Ƃ������B
���i�̓����F��2���}�܂Ŕ��B�B��1�E��2���}�͊�{�I�ɉ~���`�ŁA��[1cm�t�߂���ׂ��Ȃ��ĉ~���`�ɂȂ�B�����̂�1�����̑�2���}�̏ꍇ�A��̒��a8mm�`1cm�A�����͎w��`�R�����{�[�X�Q�̂�1.5�`3cm�A�ő��R�����{�[�X��`�ő���Q�̂�7.5cm�قǂ܂ŐL�т�B�אڂ�����2���}�̒����̊Ԃ̋����́A�O�҂�1�`2cm�قǂ�����҂ł͂���ȏ�ɂȂ�B��1���}�Ƒ�2���}�̂Ȃ��p�́A�O�҂ł�50�����邪�A��҂ł�40�������B�����̂͑��E���ŁA�O�a��3�`5mm�A��[�����R�łقƂ�Ǔˏo���Ȃ��B1���u�ǂ͂悭���B���A����1R�ɒB����B���ˌ̂͊O�a2�`3mm�̂��̂������A��ɕ@��`���O����������O�ُ�ŁA�J�����͉~�`�`���`�B1���u�ǂ͒���2/3�`1R�B�����́A���ˌ̂̑��ʂł͘]��A����ȊO�ł͖Ԗڏ�ŁA�\�ʂɂ͒P���[���⑤�Ξ������B����B
�������F���Օ��I�Ȋ�ʈ�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�`���m���匎�B��q���ł͋L�^����Ă��Ȃ��B
�⑫�F�{���Acropora glauca �i�J���r�~�h���C�V��A. pruinosa �q���G�_�~�h���C�V�ƍ�������Ă���A�����ł̐����͍Č�������K�v������B�쑺�E�ڍ�(2005)��Acropora aff.samoensis �͖{��B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FA. excelsa�Ƃ��ċL�^�����\������

�������ʐ^�̌Q�̂Ƃ͕ʌQ��(���������n�_�̓����[�ō̏W���ꂽ����)
|
|
| |
Acropora valida (Dana, 1846)
�z�\�G�_�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Acropora Oken, 1815 �~�h���C�V�� |
|

�B�e�F��@��(����q���n���̊≮�A���[2m)

|

��2���}�̒��a��7�`9mm�B
|
|
�����`�F�R�����{�[�X��A�ő��R�����{�[�X��A�ő���Q�́B
��̕��̐F�ʂƓ����F�W�ΐF�`�W���F�B�}�̐�[�����F�ɂȂ邱�Ƃ�����B
���i�̓����F��2���}�܂Ŕ��B�B��2���}�͉~���`�ŁA�����̂�1�����̏ꍇ�A��Œ��a7�`9mm�A����3�`5cm�B�אڂ�����2���}�̒����̊Ԃ̋�����1.5�`2cm�قǂł悭�����B�����̂��k���ŁA�O�a2.5mm�O��A1�`2mm�قǓˏo����B1���u�ǂ͒���1/2�`2/3R�B���ˌ̂͊Ǐ�`�@��ŁA�J�����͉~�`�`���`�B�O�a1�`2mm�ŁA�悭�ˏo������̂Ƃ��Ȃ����̂�2�^�C�v�����݂���B1���u�ǂ̔��B�������A����1/3R�����B�����͎�ɖԖڏ�ŁA���ˌ̂ł͑��ʂ��]��ɂȂ邱�Ƃ�����B�����\�ʂɂ͒P���[���⑤�Ξ������B����B
�������F���ʂ��̂悢�ʒr�E�ʌ�A�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Acropora glauca �i�J���r�~�h���C�V�ƍ�������Ă��邽�߁A��q���Ȗk�ł̐����͍Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Alveopora japonica Eguchi, 1975
�j�z���A���T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Alveopora de Blainville, 1830 �A���T���S�� |
|

�B�e�F�k��T�q(���茧�Δn�s���m�Y�A���[3m)

|

䰌a��3�`4mm�̂��̂������B
|
|
�����`�F���Q�̂ŁA���ꂢ�ȋ���ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F��Ɋ��F�`�Ê��F�ŁA���ƐG���[�͔������Ƃ��قƂ�ǁB�G��̐�[�͊ۂ��A�����̑傫���͂ǂ���قړ����B
���i�̓����F�̂͑��p�`�`���p�����~�`�B���Q�̓��ł̌̂̑傫���͂��s�����ŁA������䰂͒��a3�`4mm�A�[��3mm�قǁB�Q�̂ɂ���ẮA����炪�Ƃ���5mm����傫�Ȍ̂��܂ނ��̂�����B�u�ǂׂ͍��j���2���܂Ŕ��B���A���ˏ�ɔz��B1���u�ǂ͒����A䰐S�߂��܂ŒB����B�����͒ʏ햢���B�����A䰉����ł�1���u�Ǔ����̗Z����A�����I�ȃV�i�v�e�B�L�����ւ̌`�����H�Ɍ�����B2���u�ǂ͒�����1���u�ǂ̔����ȉ��ŁA������1���u�ǂƓ����x�����A�ꕔ�������B�̏ꍇ������B䰕ǂ͔����A���E���ł��낢�B䰕ǂɊJ�����E�͏c���̑ȉ~�`�`�~�`�ŁA��r�I�K���������㉺���E�ɕ���Ō�����B䰕Ǐ㉏�ɂ́A��[���G���ȋ������悭���B����B
�������F��ʈ�̐��[15m�Ȑ���p�̑������܂Ō����A���p���ł͑�Q�����`�����邱�Ƃ�����B
�����ł̕��z�F��t�����Y�E�������B��`����s���B�����EVeron (1995) �ł͎�q���ŋL�^����Ă��邪�A����܂ł̉�X�̒����ł͊m�F�ł��Ă��Ȃ��B
�⑫�F�{��̌��L�ڂ͍]��(1968)�ł͂Ȃ��]��(1965)�B�^�C�v�Y�n�����{�ł��邱�Ƃ͊ԈႢ�Ȃ����A�c�O�Ȃ��ƂɃ^�C�v�W�{���w�肳��Ă��Ȃ����߁A���m�ȃ^�C�v�Y�n�͖���̂܂܂ɂȂ��Ă���B�{��́A��q���Ȗk�ł�Alveopora spongiosa �A�����L�T���S�ƍ�������Ă���\�������邽�߁A�����ł̐����ɂ��Ă͍Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
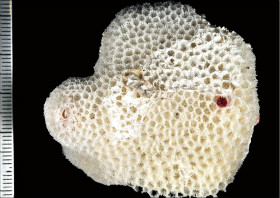
Alveopora spongiosa Dana, 1846
�A�����L�T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Alveopora de Blainville, 1830 �A���T���S�� |
|

�B�e�F������^(����q���n���̊≮�A���[14m)
|

䰌a�͏�����2mm�O��B
|
|
�����`�F��ɔ핢��`���Q�̂ŁA�Q�̂��������Ƃ��͋���ɂȂ�₷���B�傫���Ȃ�ƁA�Q�̕\�ʂɕ����̉~����ˋN���`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA���̎��ӂƐG��̐�[�������Ȃ�B�|���v�S�̂������ۂ��Q�̂�����B�G�肪�L�тĂ��Ȃ��|���v�͒���̉ԕُ�Ɍ�����B
���i�̓����F�̂͑��p�`�`�~�`�ŁA䰌a�Ɛ[����2mm�O��B�u�ǂ͐j���2���܂Ŕ��B����B1���u�ǂ�䰉����̂��̂قǒ����A䰐S�܂��͂��̎�O�܂ŐL�т�B2���u�ǂ͔��ɒZ���čׂ��A����ł͊m�F���Â炢�B�����͒ʏ햢���B�����A䰉����ł�1���u�ǂ̓����̗Z����A�����I�ȃV�i�v�e�B�L�����ւ̌`�����H�Ɍ�����B䰕ǂ͔������E���������ŁB�E�͏c���̑ȉ~�`�`�~�`�ŁA�傫������ѕ��͕s�K���B䰕Ǐ㉏�ɂ͐�[����������_��̋��������B����B
�������F�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ[�Ō�����B�J���I�Șp�ł́A��������Ƃ���Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�B��q���ł͋H�B
�⑫�F�{��́A䰌a�������x��Alveopora excelsa �AA. tizardi ��A. fenestrata �i�����3��͑S�Ė{�K�C�h���f�ڎ�j�ƍ�������Ă���B�܂��A�����ȋ���ŐG�肪�����ۂ��Q�̂́AA. japonica �j�z���A���T���S�Ƃ��Ԉ���₷���B����A�����̎�̍����ł̏ڍׂȕ��z�������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FAcropora cuneata�Ƃ��ċL�^�����\������
|
|
| |
Astreopora macrostoma Veron and Wallace, 1984
�I�I�N�`�A�i�T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Astreopora de Blainville, 1830 �A�i�T���S�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[5m)

|

䰌a��3mm�O��B
|
|
�����`�F�핢��`�����Q�́B����������o���Č�����ɂȂ�A�V���������ʂɎx����̐��ꉺ���蕔���s�K���Ɍ`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�W���F��ΐF�B
���i�̓����F�̂́A�����\�ʂ���������3mm�قǓˏo������̂���A�قږ��݂��Ă�����̂܂ŗl�X�B�Q�̒��S�t�߂̌̂͌������s�K�������A�Q�̎����ł͉����������Ƃ������B�אڂ���̂̊Ԋu�͕s�����B䰂͉~�`�`���ȉ~�`�ŁA���a3mm�O��B�u�ǂ͒ʏ�2���܂Ŕ��B���A䰕Ǐ㉏�ł킸���ɓˏo����B1���u�ǂ͔�ŁA䰕Ǐ㉏�͔��ɒZ�����A䰉����͒����A䰐S�߂��܂ŐL�т�B1���u�ǂ̒��ɂ́A�������p�Ȃ��ėאڂ���1���u�ǂɗZ���������̂�������B2���u�ǂ͔�A�H�ɐj��ŁA���ɒZ���A䰕Ǐ㉏����䰉����܂łقړ��������B�����͑��E���ŁA�\�ʂɂ͑����Ē����ו���[�������W����B�ˏo�����̂�䰕NJO���ł́A�����u�ǂ̉������ɕ��ˏ�ɕ��сA���̈ꕔ�͔�ɗZ������1���E2���]���`�����邱�Ƃ�����B
�������F�Օ��I�Șp�A�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʈ�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Astreopora gracilis �Z���x�C�A�i�T���S�i�{�K�C�h���f�ڎ�j��A. incrustans �Ȃǂƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Astreopora incrustans Bernard, 1896
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Astreopora de Blainville, 1830 �A�i�T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Óc�A���[10m)

|

䰌a��2mm�����ŏ������B
|
|
�����`�F�핢��Q�́B
��̕��̐F�ʂƓ����F���F�`�W���F��ΐF�B
���i�̓����F�̂́A�Q�̕\�ʂ���1�`1.5mm�قǓˏo���A�̂̊Ԋu������͕s�K���B䰂͉~�`�A䰌a2mm�����ŏ������B�u�ǂ�2���܂Ŕ��B�B1���u�ǂ͔�ŁA䰕Ǐ㉏�ł͔��ɒZ�����A䰐S�߂��ł͒����Ȃ�B䰐S�߂��܂ŒB����1���u�ǂ̒��ɂ́A���������p�Ȃ��ėאڂ���1���u�ǂɗZ��������̂�����B2���u�ǂ͔��ɒZ���A䰕Ǐ㉏����䰓��̉����܂łقړ��������B��ɂȂ炸�ɐj��ɂȂ�u�ǂ�����B�����͑��E���ŁA���̕\�ʂɂׂ͍��Z���P���[����ו���[�������B����B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�g�Q�̉e�������Ȃ��Ƃ���ł́A���Ō����邱�Ƃ�����B
�����ł̕��z�F���m�������ȓ�B
�⑫�F�{���䰌a�̏�����Astreopora myriophthalma �A�i�T���S�AA. expansa �C�^�A�i�T���S��A. listeri �q���A�i�T���S�i�����3��͑S�Ė{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���B����āA�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
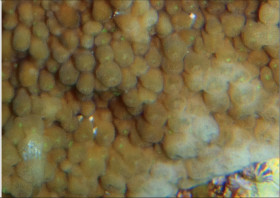
Isopora aff. cuneata (Dana, 1846)
�q���j�I�E�~�h���C�V |
Acroporidae Verrill, 1902 �~�h���C�V��
Isopora Studer, 1878 �j�I�E�~�h���C�V�� |
|

�B�e�F�[���T�L(���V�\�s�匴�A7m)
|

�����͔̂��ʂ��ɂ����B
|
|
�����`�F�핢��Q�́B�Q�̕\�ʂ��R�u��ɕs�K���ɐ���オ��B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA�N���[���F�ɂȂ���̂�����B
���i�̓����F�^��܂��͎ߏ�������������̒����̂ƕ��ˌ̂��A�Q�̕\�ʂɖ��W���ĕ��z����B�����̂͊Ǐ�ŁA�O�ǂ������k���A�O�a2�`3mm�A1�`3mm�قǓˏo����B1���u�ǂ̒���1/2�`2/3R�B���ˌ̂ɂ́A�O�a2mm�O��̊Ǐ�`�����Ǐ�̂��̂ƁA�O�a1mm�قǂłقږ��݂����Ǐ�̂��̂�2�^�C�v�����݂���B�O�҂̊Ǐ�̂�1�`2mm���x�ˏo����B1���u�ǂ̒����͂ǂ��1/2R�܂ŁB�����͑e�����łȖԖڏ�ŁA�\�ʂɂ͍ו���[�������B����B�܂��ˏo�������ˌ̂̑��ʂł́A�ו���[���̈ꕔ���Z�����Ď֍s�ו���[���ɂȂ邱�Ƃ�����B
�������F�g������̔��ɋ����ʉ��`�ʎΖʁA��ʎΖʂ̊�Տ�⋐�I��Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�{���Isopora palifera �j�I�E�~�h���C�V�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B�{��́A�Q�̌`����ˌ̂̌`��I. cuneata �i�{�K�C�h���f�ڎ�j�Ƃ͑S���قȂ��Ă���B
* ��q�����L�^��

|
|
| |
Montipora aequituberculata Bernard, 1897
�`�a�~�E�X�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|
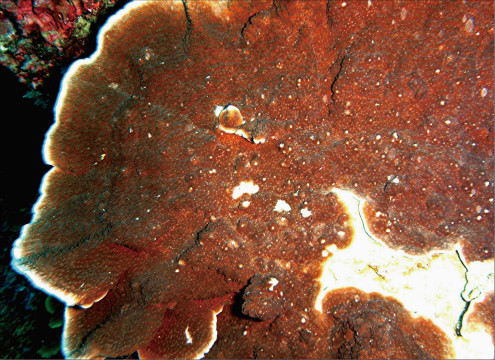
�B�e�F�쑺�b��(����q���n���̊≮�A���[5m)

|
䰌a��0.5�`0.6mm�B
|
|
�����`�F���̔�����B�Q�̂̒��a�͖�40cm�B
��̕��̐F�ʂƓ����F�|���v�E�����͊��F�A�Q�̎����͖��F�B
���i�̓����F�͔̂�Q�̂̏㉺���ʂɕ��z���A���ʂ̌̂͏�ʂ̂��̂����a��ɕ��z��������(䰌a0.3�`0.4mm)�B��ʂł͌̂͑a��ŕs�ψ�ɕ��z���A�̊Ԋu�͌�1�`5���ł���B�̂͋����ɖ��v������̂ƁA�킸���ɓˏo������̂Ƃ�����B䰌a��0.5�`0.6mm�B�u�ǂ͞���̗���Ȃ�A�T���ĕs���S�E�s�K���ł���B�����u�ǂ͌̂ɂ���ĕБ�1�{�݂̂��F�߂��A�����͖�0.8R�B1���u�ǂ͒���0.7R�ȉ��A2���u�ǂ͒���0.4R�ȉ��ŁA���҂̒����͊T���Ė��ĂɈقȂ邪�A�̂ɂ���Ă͈������ɑ����ꍇ������B䰕Ǘւ͖��ĂŃ����O����Ȃ��B������ɂ͔����ˋN����▧�ɕ��z���A�傫����`�̓o���c�L���傫���B�����ˋN�͓��Ɍ̂̎��͂Ŕ��B���A�ˋN���ڍ����Č̂���芪���ꍇ������B�Q�̎����͎̌̂��������Ɍ������ČX���X��������A���̂悤�Ȍ̂ł͌̌���̔����ˋN�Q���̂�키�悤�ɑO��(��������)�Ɍ������ĐL������B
�������F�ʊO���̐B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F������Q�̂��`������Montipora foliosa �E�X�R�����T���S�i�{�K�C�h���f�ڎ�j�Ɏ��邪�A���̎�͒�������ˋN�������Ƃŋ�ʂ����B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Montipora aff. conferta Nemenzo, 1967
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[7m)

|

䰌a�͖�0.6mm�B
|
|
�����`�F�핢��܂��͏����B�Q�̂̒��a�͖�30cm�B
��̕��̐F�ʂƓ����F�|���v�E�����͊��F�A����ˋN�̕\�ʂ̋����͖��F�B
���i�̓����F������ɂ́A�`���s�K���Ȓ��a1�`2mm�̗���ˋN����▧�ɕ��z���A�����ˋN�������B�͖̂��ɕ��z���A�̊Ԋu�͌�1���ȓ��ł���B�̂͋������ɖ��v���A�ˏo���Ȃ��B䰌a�͖�0.6mm�B�u�ǂ͂��Z���A�ŏ���͂�����ɘp�Ȃ���B�����u�ǂ�1�܂��͕Б�1�����F�߂��A�����͖�0.7R�B1���u�ǂ͊��S�E�s�K���Œ���0.5R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ���0.4R�ȉ��ŁA�����I��1���E2���u�ǂ��������ɑ����ꍇ������B䰕Ǘւ͖��ĂŁA䰕Ǘւ̊O���ɂ͗��n�т��F�߂���B�����͑e���B�̂̎��͂𗱏�ˋN���s���S�ȋ����Ǘl�Ɏ��͂ޏꍇ������B
�������F���[10m�O��̐B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�FMontipora conferta �i�{�K�C�h���f�ڎ�j�Ɏ��邪�A���̎�̊u�ǂ͖{������Z���A�܂��A䰕Ǘւ��s���Ăŗ��n�т��������Ƃŋ�ʂ����B
* ��q�����L�^��

|
|
| |
Montipora confusa Nemenzo, 1967
�~�_���A�~���R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Y�A���[10m)
|
䰌a��0.8�`0.9mm�B
|
|
�����`�F�핢��̊�Ղ���w��܂��͒���̕s�K���ŒZ����^�ˋN������ɐL���B��Վ����̈ꕔ�͔�ɒ���o���B�Q�̂̒��a�͖�60cm�B
��̕��̐F�ʂƓ����F�����͒W���F�A�|���v�͊��F�B
���i�̓����F�Q�̕\�ʂ͗t���l�̖ԖڏN�ɔ���邪�A���N�̑�����`�͕s�K���ł���B�̂͗��N�Ԃɕ��сA�������ɖ��v���Ă��ēˏo���Ȃ��B䰌a��0.8�`0.9mm�B�u�ǂ͞���̗���Ȃ�A�T���ĕs���S�E�s�K���B�����u�ǂ͌̂ɂ���ĕБ�1�{�̂ݔF�߂��A������0.7�`0.8R�B1���u�ǂ͒���0.7R�ȉ��A2���u�ǂ�����0.4R�ȉ��ŁA���҂̒����͖��ĂɈقȂ�B䰕Ǘւ͖��ĂŁA�����Ă����������O����Ȃ����A�h�[�i�c��ɐ���オ��ꍇ������B�����͔����ˋN�������B
�������F���[10m�t�߂̐B
�����ł̕��z�F��q���݂̂���m����B��q���ł͋H�B
�⑫�FMontipora sp. AMIME. �A�~���R�����T���S�Ɏ��邪�A���̎�̖ԖڏN�ׂ͍���r�I�ψ�ɕ��z���邱�Ƃƒ���ˋN�������Ȃ����ƂŖ{��Ƌ�ʂ����B�V�̘a���͖ԖڏN���s�K���Ȃ��ƂɈ��ށB
* ��q�����L�^��

|
|
| |
Montipora danae Milne Edwards and Haime, 1851
�f�[�i�C�{�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Y�A���[7m)
|

䰌a�͖�0.8mm�B
|
|
�����`�F�핢��B�Q�̂̒��a�͖�50cm�B
��̕��̐F�ʂƓ����F�����E�|���v���ɒW���F�B
���i�̓����F�Q�̕\�ʂɂ͌`��傫��(���a2mm�O��)���قڋψ���n��ˋN���������邪�A�Q�̎����ł݂͌��ɐڍ����ĒZ������ˋN���`������B�̂��n��ˋN�Ԃ݂̂ɕ��z���A�̊Ԋu�͌�1���ȓ��ŁA�������ɖ��v���ˏo���Ȃ��B䰌a�͖�0.8mm�B�u�ǂ͞���̗���Ȃ�A�����u�ǂ͕s���āA1���u�ǂ͊��S�E���s�K���Œ�����0.6R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ�����0.4R�ȉ��ł���B����̊u�ǂ͂��Z�����A䰂����ڂ܂邽��䰒�ł�1���u�ǂ����S�܂ŐL�т�䰒ꕔ���ǂ��B䰕Ǘւ͖��ĂŁA�ׂ������O����Ȃ��B�����͔����ˋN�������A���ׂ͍��Z���B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȓ�B��q���ł͂��H�B
�⑫�FMontipora verrucosa �C�{�R�����T���S�ɍ������邪�A���̎��䰌a�͖�1mm�Ƒ傫�����ƁA�u�ǂ��Z����䰕ǂ����ڂ܂炸�ɐ����ɗ������ނ���䰓��̓|�c���ƌ����͂�����Č����邱�ƁA�Q�̎����Ő���ˋN���`�����Ȃ����ƂȂǂ̓����Ŗ{��Ƌ�ʂ����B
* ��q�����L�^��

|
|
| |
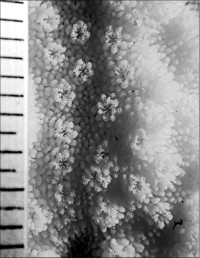
Montipora aff. digitata (Dana, 1846) sp. 3
�i�K�G�_�R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[2m)
|

䰌a�͖�0.4mm�B
|
|
�����`�F���}��B�Q�̂̍�����10cm�A��̎}�̑�����1cm�B
��̕��̐F�ʂƓ����F�����͒W���F�A�|���v�͖��F�B
���i�̓����F�̂͋ψ�ɖ��ɕ��z���A�̊Ԋu�̖͌�1���ł���B�̂͋������ɖ��v���ˏo���Ȃ��B䰌a�͖�0.4mm�B�����u�ǂ��܂�1���u�ǂ͂�����ɓˏo���A1���u�ǂ�2���u�ǂ͒��������ĂɈقȂ�B�����u�ǂ͊�{�I��1�{���F�߂��A������0.8�`1.0R�B1���u�ǂ͊��S�E���s�K���Œ�����0.7R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ�����0.3R�ȉ��ł���B�̂ɂ���ẮA䰕Ǘւ����ĂŃh�[�i�c��ɐ���オ��B�����͔����ˋN�������A���͑@�ׂŒZ���A�����ǂ̔��B���ア���߁A�����\�ʂ͊��炩�Ɍ�����B
�������F�ʒr���̐B
�����ł̕��z�F��q���ȓ�B
�⑫�F�����ǂ�����ˋN�����������\�ʂ����炩�Ȃ��ƂƁA�����u�ǂ�1���u�ǂ��������Ƃő��̎��}��ɂȂ�R�����T���S�ނƋ�ʂ����B�V�̘a���͋߉���Montipora digitata �G�_�R�����T���S�i�{�K�C�h���f�ڎ�j�����u�ǂ��������ƂɈ��ށB
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FM. informis�Ƃ��ċL�^�����\������
|
|
| |
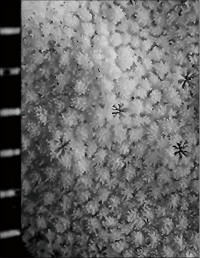
Montipora grisea Bernard, 1897
�O���Z�A�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[6m)

|
䰌a�͖�0.7mm�B
|
|
�����`�F�핢��B�Q�̂̒��a�͖�30cm�B
��̕��̐F�ʂƓ����F�����͒W���F�B
���i�̓����F�͕̂s�K���ɕ��z���A�̊Ԋu�͌�1�`3���ł���B�قƂ�ǂ̌̂͋�������1mm���x�ˏo����B䰌a�͖�0.7mm�B�u�ǂ͞���̗���Ȃ邪�A�����u�ǂ͌̂ɂ�莕����`������ꍇ������B�܂��A�����u�ǂ͊T���ĕs���Ăł��邪�A�̂ɂ���Ă͕Б�1�{���݂̂��F�߂��A�����͖�0.7R�ł���B1���u�ǂ͊��S�E���s�K���Œ�����0.6R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ�����0.4R�ȉ��B䰕Ǘւ͂�▾�ĂŁA���������O��B������ɂ̓R���p�N�g�Ȕ����ˋN�����ɕ��z���A�̂̎��͂̂��̂�䰕ǗւƐڍ����ĕs���S�ȓ����䰍��ǂ��`������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�̂̂قƂ�ǂ��킸���ɓˏo����̂��{��̓����ŁA�����ˋN�������̎�Ƌ�ʂ����B
* ��q�����L�^��

|
|
| |
Montipora aff. hispida (Dana, 1846)
�z���h�g�Q�R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Y�A���[5m)

|

䰌a��0.6�`0.7mm�B
|
|
�����`�F��v�����͔핢��ŁA�����͕����I�ɔ�ɒ���o���B�܂��A����Ɍ������ĕs�K���Ȓ���ˋN��L���ꍇ������B�Q�̂̒��a�͖�40cm�B
��̕��̐F�ʂƓ����F�����͊��F�E�P���F�E�P�Ί��F�E�P�����F�ƕψق�����A�|���v�͋����Ɠ��n�F�ƂȂ�ꍇ�������B
���i�̓����F�̂̕��z�͕s�ψ�ŁA��^�ˋN��ł͑a��A�ˋN�Ԃł͖��W����X��������B�͖̂��v�^�Ɠˏo�^��2�^������B䰌a��0.6�`0.7mm�B�����u�ǂ͂��s���ĂŁA����0.5�`0.6R�A�T����1�{�݂̂����ʂ���邪�A1��䰉��Őڍ�����ꍇ������B1���E2���u�ǂ͂قڊ��S�E�K���I�ŁA�������A����0.5R�ȉ��B䰕Ǘւ͖��ĂŁA�����O��܂��͊Ǐ�ŁA��҂̏ꍇ�͌̂����Ăɓˏo����B������ɂ͔����ˋN���s�ψ�ɕ��z���A�傫����`�̃o���c�L���傫�����A�`�����P���ŕ������A�\�ʂ̞��̖ё����ג����X��������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F�a�̎R�����{�`��q���B��q���ł͂��H�B
�⑫�FMontipora hispida �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�Ƃ͊u�ǂ�����ˋN�̌`�ɑ��Ⴊ�F�߂��A�ʎ�Ɣ��f�����B�{��͎�q���Ȗk�̍��ܓx�C��ɕ��z����ŗL��Ǝv���A�V�̘a��������Ɉ��ށB
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Montipora cf. informis Bernard, 1897
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[5m)

|
䰌a��0.6�`0.7mm�B
|
|
�����`�F�핢��B�Q�̕\�ʂ͕������s���Ăȑ召��ᎏ�ˋN���U�݁B�Q�̂̒��a�͖�40cm�B
��̕��̐F�ʂƓ����F�����͊��F�B
���i�̓����F�̂͑a��ɁA���s�ψ�ɕ��z���A�̊Ԋu�͌�1�`4���ł���B�̂̑����͋����ɖ��v���邪�A�ꕔ�͂킸���ɓˏo����B䰌a��0.6�`0.7mm�B�u�ǂ͊�{�I�ɞ���̗���Ȃ邪�A�����u�ǂ�1���u�ǂ̈ꕔ�͎�����`������ꍇ������B1���u�ǂ�2���u�ǂ͖��Ăɒ������قȂ�B�����u�ǂ�1���F�߂��A�����͖�0.6�`0.8R�ł���B1���u�ǂ͊��S�E�K���I�Œ�����0.7R�ȉ��A2���u�ǂ͕s�K���E�s���S�Œ�����0.4R�ȉ��ŁA�T���Ĕ��B�������B䰕Ǘւ͖��ĂŃ����O��B�����ˋN�͔w���Ⴍ�A�T���ăR���p�N�g�ŁA�ψ�ɖ�������B�܂��A�傫����`���قڋψ�ł��邪�A�̂ɂ���Ă�䰕ǗւƐڍ������s���S�ȊǏ�̋����ǂ��`������ꍇ������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�FMontipora informis �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�ɂ悭���邪�A�Q�̌^��u�ǂ̍\���ɑ��Ⴊ�F�߂���B�����ˋN�������̎�Ƃ́A�{��̔����ˋN�͒Z���Ăقڈ�l�ɖ������邱�ƁA1���u�ǂ���������2���u�ǂ������B�Ȃ��ƁA�̂��a��ɕ��z���邱�Ƃ̓����ŋ�ʂ����B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Montipora millepora Crossland, 1952
�~���|���R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[10m)

|

䰌a��0.4�`0.5mm�B
|
|
�����`�F�핢��B�Q�̕\�ʂ͕������召�s�K����ᎏ�ˋN���U�݂���B�Q�̂̒��a�͖�30cm�B
��̕��̐F�ʂƓ����F�����͒W�����F�E�W�Ί��F�E�W�����F���A�|���v�͊��F�A�ΐF���ψق������B
���i�̓����F�̂͑a��ɕs�ψ�ɕ��z���A�̊Ԋu�͌�1�`5���ł���B�̂͋������ɖ��v������̂ƁA���ˏo������̂Ƃ�����B䰌a��0.4�`0.5mm�B�u�ǂ͞���̗���Ȃ邪�A�����u�ǂ͎�����`������ꍇ������B�u�ǂ͊����˂���A�܂��A����ɂ��ˏo����X��������B�����u�ǂ�1���܂���1���F�߂��A������0.8R�B1���u�ǁE2���u�Nj��ɕs���S�E�s�K���ŁA1���u�ǂ̒�����0.7R�ȉ��A2���u�ǂ̒���0.5R�ȉ��ŁA�̂ɂ���Ă͗��҂̒������������ɑ����ꍇ������B�ˏo�^�̌̂ł�䰕Ǘւ͖��ĂŁA䰕Ǘ֏�₻�̎��̞͂��Ƌ��ɒႭ����オ��B�����͔����ˋN�������A���͑@�ׂŒZ���A�����\�ʂ͊��炩�Ɍ�����B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂��H�B
�⑫�FMontipora mollis �����X�R�����T���S��M. aff.spongodes �~�_���C�{�R�����T���S�Ɏ���B�O�҂͌̂����ɕ��z����������ˏo���Ȃ����ƂŁA��҂��n��ˋN�������ƂŁA�{��Ƌ�ʂ����B
* ��q�����L�^��

|
|
| |
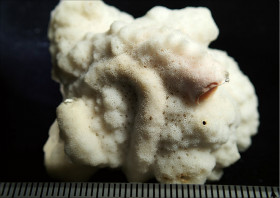
Montipora mollis Bernard, 1897
�����X�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[10m)
|
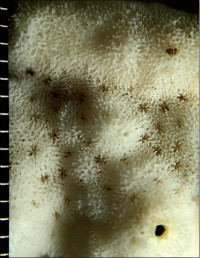
䰌a��0.6�`0.7mm�B
|
|
�����`�F�핢��`�����B�Q�̂̒��a�͖�50cm�B
��̕��̐F�ʂƓ����F�����E�|���v���ɁA���F�n�ƗΐF�n��2�^������B
���i�̓����F�Q�̕\�ʂɂ͑傫����`���s�ψ��ᎏ�ˋN���U�݂���B�͖̂��ɕ��z���A�̊Ԋu�͌�1���ȓ��B�̂͋������ɖ��v���ˏo���Ȃ��B䰌a��0.6�`0.7mm�B�u�ǂ͞���̗���Ȃ邪�A�����u�ǂ͎�����`������ꍇ������B�����u�ǂ͒ʏ�1���F�߂��A����0.9�`1.0R�A����ɂ��ˏo����B1���u�ǂ͊��S�E�K���I�Œ���0.7R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ���0.3R�ȉ��ł���B䰕Ǘւ͖��ĂŁA���n�т������B�����͔����ˋN�������A�����ǂ͊T���ĕs���Ăł���B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂�╁�ʁB
�⑫�F�O����Montipora millepora �~���|���R�����T���S�AM. aff. spongodes �~�_���C�{�R�����T���S�AM. aff.turgescens �A�o�^�R�����T���S�Ɏ���B�O�҂͌̂��a��ɕ��z���Ⴍ�ˏo���邱�ƂŁA���҂��n��ˋN�������ƂŁA��҂͊u�ǂ��Z�����n�т����邱�ƂŁA�{��Ƌ�ʂ����B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FM. spongodes�Ƃ��ċL�^�����\������
|
|
| |
Montipora monasteriata (Forskål, 1775)
�g�Q�N�{�~�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(����q���剖���A���[2m)

|
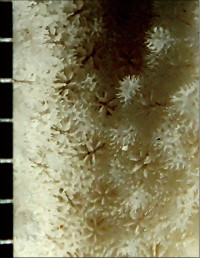
䰌a�͖�0.7�`0.8mm�B
|
|
�����`�F�핢��`�����B�Q�̂̒��a�͖�50cm�B
��̕��̐F�ʂƓ����F�|���v�E�����Ƃ��Ɋ��F���A�����͒W�����F�Ń|���v�͖��F�B
���i�̓����F�Q�̕\�ʂɂ͒��a1cm�O���ᎏ�ˋN�����ɕ��z����B�͖̂��ɕ��z���A����ᎏ�ˋN�Ԃł͌����ł���B�̂͋������ɖ��v���A�ˏo���Ȃ��B䰌a�͖�0.7�`0.8mm�ł���B�u�ǂ͒����A�����u�ǂ�1���u�ǂ͏���ɓˏo���A������`������B�����u�ǂ�1���܂���1�A������0.9R�ȉ��ŁA����1���u�ǂ��������B1���u�ǂ͊��S�E�K���I�Œ���0.8R�ȉ��A2���u�ǂ͕s���S�E�s�K���A����0.3R�ȉ��ŁA���҂̒����͖��ĂɈقȂ�Bᎏ�ˋN��ł͋����ǂ͂�┭�B����X��������A���̂悤�ȏꏊ�ł�䰕ǗւƗ��n�т����ĂɔF�߂���B������ɂ͔����ˋN���a��ɕ��z���A�ڍ����������ˋN���̂����͂ޏꍇ������B
�������F���[10m�O��̐B
�����ł̕��z�F��q���ȓ�B��q���ł͕��ʁB
�⑫�FMontipora aff.turgescens �A�o�^�R�����T���S�ɍ������邪�A���̎�͊u�ǂ��Z���A�܂��A�����ˋN���������ƂŖ{��Ƌ�ʂ����B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FM. turgescens�Ƃ��ċL�^�����\������
|
|
| |
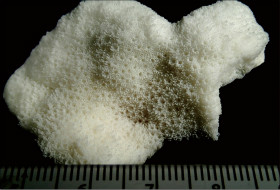
Montipora peltiformis Bernard, 1897
�����T�L�R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Y�A���[7m)

|

䰌a��0.7�`0.8mm�B
|
|
�����`�F�핢��A�\�ʂɂ͕s�K����ᎏ�ˋN���U�݁B�Q�̂̒��a�͍ő�Ŗ�60cm�B
��̕��̐F�ʂƓ����F�����͊��F�A�|���v�͎��F�܂��͊��F�B
���i�̓����F�Q�̎����������Č͖̂��W���ĕ��z���A�̊Ԋu�͌�1���ȓ��ł���B�̂͋����ɖ��v���邪�A�킸���ɓˏo����ꍇ������B䰌a��0.7�`0.8mm�B�u�ǂ͞���̗���Ȃ邪�A�̂ɂ������u�ǂ�������`������ꍇ������B�����u�ǂ͊T���ĕs���Ăł��邪�A�Б�1���݂̂��F�߂���ꍇ������A�����͖�0.8R�ł���B1���u�ǂ͊��S�E�s�K���Œ�����0.5R�ȉ��A2���u�ǂ͕s���S�E�s�K���A������0.4R�ȉ��ŁA�̂ɂ��1���E2���u�ǂ��������ɑ����ꍇ������B䰕Ǘւ͖��Ăȃ����O��B������ɂ͂��R���p�N�g�Ȕ����ˋN���s�ψ�ɕ��z���A�̂̎��͂ł킸���ɔ��B����X��������B�܂��A�̎��͂̔����ˋN���m���ڍ����Č̂�s���S�Ɏ��͂ޏꍇ������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂�╁�ʁB
�⑫�F�{��͌̂����W���ċψ�ɕ��z���邱�ƁA�����ˋN�͂��R���p�N�g�ŕs�ψ�ɕ��z���邱�ƁA�����ˋN���ڍ����čג����V����̐���ˋN���`�����Ȃ����Ɠ��̓����ŁA�����ˋN�������̎�Ƌ�ʂ����B�{��̓V����̐���ˋN�����Ƃ��ăC�^�C�{�R�����T���S�̘a�����^����ꂽ��(����E����, 1985)�A���̘a���͌듯��Ɋ�Â��B�V�̘a���͖{�킪���F�̃|���v�������̂��������ƂɈ��ށB
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FM. undata�Ƃ��ċL�^�����\������
|
|
| |
Montipora aff. spongodes Bernard, 1897
�~�_���C�{�R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s���Y�A���[10m)

|
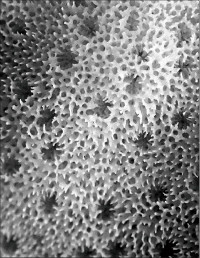
䰌a�͖�0.6�`0.7mm�B
|
|
�����`�F�핢��B�Q�̂̒��a�͖�30cm�B
��̕��̐F�ʂƓ����F�Q�̎����Ɨ���ˋN�̋����͒W�����F�A���̑��̋����ƃ|���v�͒W���F�B
���i�̓����F�Q�̕\�ʂɂ͑傫����`���ɂ߂ĕs�ψ���n��ˋN���a��ɕ��z����B�͕̂s�ψ�ɕ��z���A�̊Ԋu�͌�1�`5���ł���B�̂͋������ɖ��v���ˏo���Ȃ��B䰌a�͖�0.6�`0.7mm�B�u�ǂ͞���̗���Ȃ�A�s���S�E�s�K���ł���B�����u�ǂ�1���܂���1���F�߂��A����1.0R�ȉ��A1���u�ǂ�0.7R�ȉ��A2���u�ǂ�0.4R�ȉ���1���u�ǂ�2���u�ǂ͌̂ɂ���Ă͈������ɑ����ꍇ������B䰕Ǘւ͂�▾�ĂŁA�h�[�i�c��ɐ���オ��ꍇ������B�����͔��ɑ@�ׂȍ��g�݂��Ȃ�A�����@�ׂŒZ�����߁A�����\�ʂ͊��炩�Ɍ�����B�����ˋN�������B
�������F���[10m�O��̐B
�����ł̕��z�F�a�̎R�����{�`��q���B��q���ł͋H�B
�⑫�F�n��ˋN�����ގ���Ɏ��邪�A�{����n��ˋN�͕s�K���ł��a��ɕ��z���邱�Ƃŋ�ʂ����BMontipora spongodes �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�ɍ��i�`�Ԃ��������邪�A���̎���n��ˋN�������Ȃ��B�Ȃ��A�{��͎�q���Ȗk�̍��ܓx�C��ɕ��z����ŗL��Ǝv����B�V�̘a���͕s�K�����n��ˋN�������ƂɈ��ށB
* ��q�����L�^��

|
|
| |
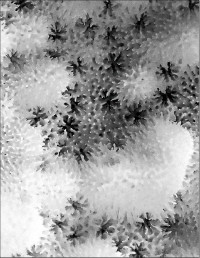
Montipora aff. turgescens Bernard, 1897
�A�o�^�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[6m)

|
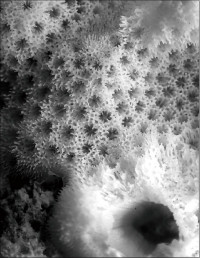
䰌a��0.6�`0.8mm�B
|
|
�����`�F�핢��`���B�Q�̂̒��a�͍ő��1m�B
��̕��̐F�ʂƓ����F�|���v�E�����ɂ͕ψق������A�W���F�A���F�A�ΐF�A�Ί��F�A�Ó��F��������B
���i�̓����F�Q�̕\�ʂɂ͑召�̕s�K����ᎏ�ˋN���U�݂���B�̂͒ʏ햧�ɕ��z���A�������ɖ��v����B䰌a��0.6�`0.8mm�B�u�ǂ̒����͌̂�Q�̂ɂ���ĕψق��傫�����A�T���ĒZ���B�ʏ�A�����u�ǂ��F�߂�꒷���͖�0.7R�A1���u�ǂ͕s���S�E�s�K���Œ���0.6R�ȉ��A2���u�ǂ͕s���S�E�s�K���A����0.3R�ȉ��ŁA�����I��1���E2���u�ǂ��������ɑ����ꍇ������B䰕Ǘւ͖��ĂŁA䰕Ǘւ̊O���ɂ͗��n�т��F�߂���B�ʏ�Aᎏ�ˋN��ł͋����ǂ���┭�B����B�����͔����ˋN�������B
�������F���[10m�ȓ��̐B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͕��ʁB
�⑫�FMontipora mollis �����X�R�����T���S��M. monasteriata �g�Q�N�{�~�R�����T���S�Ɏ���B�O�҂͗��n�т��������ƂŁA��҂͋�����ɔ����ˋN�������ƂŁA���ꂼ��{��Ƌ�ʂ����B�܂��A�{���M. turgescens �z���A�o�^�R�����T���S�i�{�K�C�h���f�ڎ�j�ɍ������邪�A���̎��䰌a�����傫���A�u�ǂ͂��Z���A�����ǂ͂�蔭�B���铙�̑��Ⴊ�F�߂���B
* ��q�����L�^��
|
|
| |
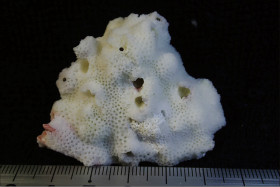
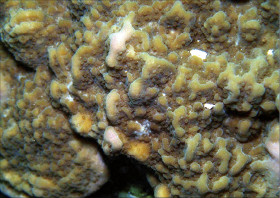
Montipora cf. undata Bernard, 1897
�a���Ȃ� |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[7m)
|
䰌a��0.9�`1.3mm�B
|
|
�����`�F�핢��`�����B�Q�̕\�ʂɂ�ᎏ�`����̒Z���ĕs�K���ȑ�^�ˋN���s�ψ�ɕ��z�B�Q�̂̍ő咷�a�͖�60cm�B
��̕��̐F�ʂƓ����F�|���v�A�����Ƃ��ɒW���F�B
���i�̓����F�Q�̕\�ʂɂ͑傫����`�̕s�K���Ȑ���ˋN���n��ˋN���s�ψ�ɕ��z����B����ˋN�͒Z���A�K���I�ȖԖڏ�\���͌`������Ȃ��B�̂͂����̓ˋN�Ԃ܂��͓ˋN���ʂɕ��z���A�ˋN��ʂɂ͕��z���Ȃ��B�̂͋������ɖ��v���邩�A�킸���ɓˏo����B䰌a��0.9�`1.3mm�ŁA�Q�̂ɂ���ăo���c�L���傫���B�����u�ǂ͂��s���ĂŁA1���u�ǂ����킸���ɒ������x�B1���u�ǂ͊��S�E�s�K���Œ���0.6R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ���0.4R�ȉ��ŁA�̂ɂ���Ă͗��҂��������ɑ����ꍇ������B䰕Ǘւ͖��ĂŃ����O��A���n�т͕s���Ăł���B�����͔����ˋN�������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȓ�B
�⑫�FMontipora undata �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{��䰌a��0.6mm�Ƃ�⏬�����A����ˋN���ג����A�Ȃ铙�A�{��Ƃ̑���_���F�߂���B�܂��A����ˋN������M. sp. AMIME. �A�~���R�����T���S��M. confusa �~�_���A�~���R�����T���S�ɂ����邪�A�{��̐���ˋN�͊�{�I�ɖԖڏ�ɂȂ�Ȃ����Ƃŋ�ʂ����B
* ��q�����L�^��
|
|
| |
Montipora aff. venosa (Ehrenberg, 1834)
�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[6m)

|

䰌a��0.7�`0.9mm�B
|
|
�����`�F�핢��`�����ŁA�Q�̕\�ʂɂ͕s�K����ᎏ�ˋN���U�݁B�Q�̂̍ő咷�a�͖�50cm�B
��̕��̐F�ʂƓ����F�����͒W���F�A�|���v�͒W���F�܂��͊��F�B
���i�̓����F�̂͂قڋψ�ɖ��ɕ��z���A�̊Ԋu�̖͌�1���ł���B�̂͋������ɖ��v���A�ˏo���Ȃ��B䰌a��0.7�`0.9mm�B�u�ǂ͞���̗���Ȃ邪�A�̂ɂ���ĕ����u�ǂȂ�т�1���u�ǂ̈ꕔ��������`������ꍇ������B�u�Ǎŏ���͕s���S�E�s�K���ŒZ�����A䰂����ڂނ��߁A�����u�ǂ�1���u�ǂ̑S�Ă܂��͈ꕔ���ꕔ�Őڍ����A�܂��A�����̌̂Ŏ��������F�߂���B�Ȃ��A�u�Ǎŏ���ł�1���u�ǂ͒���0.5R�ȉ��A2���u�ǂ͒���0.3R�ȉ��ŁA�����u�ǂ͂��s���āB䰕Ǘւ͖��Ăōׂ������O��ŁA���n�т������B�T���ċ����ǂ̔��B�͈������Aᎏ�ˋN��ł͕����I�ɔ��B����B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F�O�d���F��E�F�{���V�����[�`��q���B
�⑫�FMontipora venosa �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�ɍ��i�`�Ԃ����邪�A���̎��䰕ǂ͂قڐ����ɗ������ނ��ƁA�����ǂ��悭���B���邱�Ɠ��̑��Ⴊ�F�߂���B�Ȃ��A�{��͎�q���Ȗk�̍��ܓx�C��ɕ��z����ŗL��Ǝv����B
* ��q�����L�^��
|
|
| |
Montipora verrucosa (Lamarck, 1816)
�C�{�R�����T���S |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[7m)

|

䰌a�͖�1.0mm�B
|
|
�����`�F�핢��B�Q�̂̒��a�͖�15cm�B
��̕��̐F�ʂƓ����F�F�ʁF�����E�|���v���ɒW�Ί��F�B
���i�̓����F�Q�̕\�ʂɂ͒��a3�`4mm���n��ˋN�������B�̂͗���ˋN�Ԃ݂̂ŁA��1�����x����ĕ��z����B�̂͋������ɖ��v���ˏo���Ȃ��B䰌a�͖�1.0mm�B�u�ǂ͞���̗���Ȃ�A�����u�ǂ͕s���āA1���u�ǂ͊��S�E���s�K���Œ���0.3R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ���0.2R�ȉ��ł���B䰕ǂ͐����ɗ������݁A���u�ǂ��Z�����߁A�����͂�����Č�����B䰒ꕔ�ł͊u�ǂ��݂��ɐڍ������������`������B䰕Ǘւ͕s���āB�����͔����ˋN�������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�FMontipora danae �f�[�i�C�{�R�����T���S�ɍ������邪�A���̎��䰌a(��0.8mm)�͖{���菬�������ƁA䰒ꕔ���u�ǂɉB��Ď��ʂ�����ƁA�Q�̎����ł͕����I���n��ˋN���ڍ����Đ���ˋN���`�����邱�ƂŖ{��Ƌ�ʂ����B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FL. explanata�Ƃ��ċL�^�����\������
|
|
| |
Montipora sp. AMIME.
�A�~���R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[7m)

|

䰌a��0.6�`0.7mm�B
|
|
�����`�F�핢��A�Q�̕\�ʂɕs�K����ᎏ�ˋN���U�݁B�Q�̂̍ő咷�a�͖�40cm�B
��̕��̐F�ʂƓ����F�����E�|���v���ɒW���F�ŁA�G��̐悪���F�ɂȂ�ꍇ������B
���i�̓����F�Q�̕\�ʂ͗t���l�ׂ̍��ȖԖڏN�ɔ���A���̗��N���Ɍ̂�1�܂��͐������ԁB�̂͋������ɖ��v���A�ˏo���Ȃ��B�Q�̎����t�߂̌̂́A�����Ɍ������ĉ������ɓ|���X��������B䰌a��0.6�`0.7mm�B�u�ǂ͞���̗���Ȃ邪�A�����u�ǂ͎�����`������B�����u�ǂ͕����I��1���܂���1���F�߂��A������0.7�`0.9R�B1���u�ǂ͊��S�E�s�K���Œ���0.5R�ȉ��A2���u�ǂ͕s���S�E�s�K���Œ���0.4R�ȉ��ŁA�T����1���E2���u�ǂ͈������ł���B䰕Ǘւ͖��āA���n�т������B���͒Z���@�ׂŁA�P���Ȑj�ג������Џ���Ȃ��B�����͔����ˋN�������B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���̑��ɂ͋{�茧�싽���݂̂���̏W����Ă���B��q���ł͂��H�B
�⑫�FMontipora confusa �~�_���A�~���R�����T���S�Ɏ��邪�A���̎�̖ԖڏN�͕s�K���ŁA����ɒZ������ˋN��L�����ƂŖ{��Ƌ�ʂ����B�Ȃ��A�{��͎�q���Ȗk�̍��ܓx�C��ɕ��z����ŗL��Ǝv����B�V�̘a���͖ԖڏN���������Ɉ��ށB
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Montipora sp. TANEGA.
�^�l�K�V�}�R�����T���S�i�V�́j |
Acroporidae Verrill, 1902 �~�h���C�V��
Montipora de Blainville, 1830 �R�����T���S�� |
|

�B�e�F�쑺�b��(���V�\�s�匴�A���[10m)

|
䰌a��0.8�`0.9mm�B
|
|
�����`�F��Ղ͔핢��܂��͏����ŁA��Ղ���͎w��Ȃ�т�ᎏ�̑�^�ˋN���s�ψ�ɓˏo����B��Ղ̎����͔�ɒ���o���B�Q�̂ɂ���Ă͒���ˋN��������B
��̕��̐F�ʂƓ����F�����E�|���v���ɗΊ��F�ŁA�G��̐�[�͖��F�ɐF���������B
���i�̓����F�̂̕��z�ɂ͑a��������A��^�ˋN��ł͂قڌ�1���Ԋu�ŋψ�ɕ��z���邪�A��^�ˋN�Ԃł͌̂͂���ɖ��W����B�܂��A��^�ˋN��ł͋����ǂ����B���邪�A�K���I�ȃZ���I�C�h�\���͎������A�����Εǂ̒��Ɍ̂������ԁB�̂͑傫���A䰌a��0.8�`0.9mm�B䰕Ǘւ͖��Ăȃ����O��A���̎��͂ɂ͗��n�т��F�߂���B�u�ǂ͕ψق��傫���A��^�ˋN�Ԃɕ��z����̂ł͊u�ǂ͂قƂ�ǔ��B�����A䰓����~����ɐ[���Č�����B��^�ˋN��̌̂ł́A�ŏ㕔�̊u�ǂ͒Z���A䰒�Ɍ������ɂ�Ē����Ȃ�A䰒ꕔ�ł͕����u�ǂ�1���u�ǂ݂͌��ɐڍ����ď����Ȏ��������`������X��������B�����͑@�ׂŃX�|���W��A�����T���ĒZ���@�ׂŁA�P���Ȑj�ג������Џ���Ȃ��B
�������F���[10m�Ȑ�̐B
�����ł̕��z�F��q���ȊO�ł͋L�^����Ă��Ȃ��B��q���ł͂��H�B
�⑫�F�����ǂ������̎�Ɏ��邪�A�u�ǂ̍\���A䰕Ǘւ₻�̎��͂̍\���A�����➙�̍\�����ɑ��Ⴊ�F�߂���B�Ȃ��A�{��͖��L�ڎ�ł���\��������B�V�̘a���͎�q�������Y�n�ł��邱�ƂɈ��ށB
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Leptoseris glabra Dinesen, 1980
�Z���x�C�T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Leptoseris Milne Edwards and Haime, 1849 �Z���x�C�T���S�� |
|

�B�e�F���n�m�V(����q���n���≮�A���[10m)

|

��E�����u�ǂ̌����̈Ⴂ�����āB
|
|
�����`�F�t��`��Q�́B�Q�̂������Ȏ��́A�R�l��ɂȂ��Ă��邱�Ƃ������B
��̕��̐F�ʂƓ����F�W���F�`���F�B�Q�̎����͔����ۂ��Ȃ�B�����ȌQ�̂ł́A���S�̂�F���ł��邱�Ƃ������B
���i�̓����F�̂͌Q�̏�ʂ݂̂ɕ��z���A�T���i�X�e���C�h�^�ɔz��B���S�̂��̂������̂͑ȉ~�`�ŁA���a3�`8mm�A�Q�̎��������������ł̌̂̑傫���͕s�����ɂȂ�₷���B�Q�̎����̌̂́A�����Ɍ������ĉ������ɌX���Ȃ���ˏo����B�����ȊO�ł́A�̂������ɑ��Ăقڕ��s(�Q�̂̌Œ����𒆐S�Ƃ��Ăقړ��S�~��)�ɔz�A�Q�̂̊��������t�߂܂Ŗ��ՂȂ��A�ߐڂ��Ȃ��番�z����B�̂̊u�ǁE�]��4���܂Ŕ��B�B1���E2���u�ǁE�]��3���E4���u�ǂ��������A����ɂ悭�ˏo����B1���E2���u�ǂ͉~����`�G���Ȕ�̎����܂ŒB���邪�A3���E4���u�ǂ͒B���Ȃ��B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�����EVeron (1995) ��Veron (2000) ��Leptoseris explanata �ƌf�ڂ���Ă���͖̂{��ŁA�Z���x���T���S�̘a����S������{��ł���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
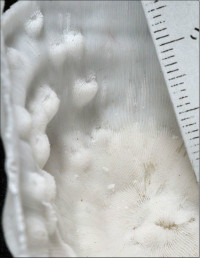
Leptoseris hawaiiensis Vaughan, 1907
�n���C�Z���x�C�T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Leptoseris Milne Edwards and Haime, 1849 �Z���x�C�T���S�� |
|

�B�e�F������^(���V�\�s��Óc�A���[17m)
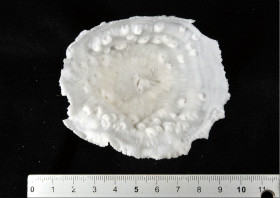
|
��E�����u�ǂ̌����̈Ⴂ���s���āB
|
|
�����`�F�t��`��Q�́B�Q�̂��܂������Ȏ��́A�R�l��ɂȂ��Ă��邱�Ƃ������B
��̕��̐F�ʂƓ����F���F�ŁA�Q�̂̎����͔����ۂ��Ȃ�B�����ȌQ�̂ł́A���S�̂�F���ł��邱�Ƃ������B
���i�̓����F�̂͌Q�̏�ʂ݂̂ɕ��z���A�T���i�X�e���C�h�^�ɔz��B���S�̂��̂������̂͑ȉ~�`�ŁA���a2�`5mm�B�Q�̏�ł̌̂̕��z�͕s�K���ŁA�قƂ�nĵ������Ȃ�����������A�����̌̂��Q�̎����ƕ��s�ɕ��ԕ���������B�̂͋����ɖ��݂�����́A�u��ɓˏo������̂���Q�̎����Ɍ������ĉ������ɌX���Ȃ���ˏo������̂ȂǗl�X�B�u�ǁE�]��4���܂Ŕ��B���A���̌�����ˏo�x���͎����ɂ�炸�قړ����B3���E4���u�ǂ�1���E2���u�ǂɔ�ׂĂ킸���ɒZ���B�̂̂����O���ł́A�]��������֍s�����肷�邱�Ƃ�����B�����͔��B�������A�������̂����Ȃ��Ȃ��B���̂��ߓ���ł́A䰐S�ɏ����Ȍ����J�����悤�Ɍ�����B
�������F�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ[�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
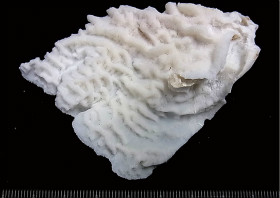
Leptoseris mycetoseroides Wells, 1954
�A�o�^�Z���x�C�T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Leptoseris Milne Edwards and Haime, 1849 �Z���x�C�T���S�� |
|

�B�e�F�쑺�b��(����q���剖���A���[2m)
|

�R�����͓��S�~��ɔ��B���₷���B
|
|
�����`�F�핢��Q�́B�Q�̕\�ʂ͋N�����������B�Q�̎����ł́A�Œ���Ղ���V�����ėt��ɂȂ��Ă��邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Q�̎�����u�ǁE�]�̏㉏�������ۂ��Ȃ邱�Ƃ�����B�����ȌQ�̂ł����S�̂͌����Ȃ��B
���i�̓����F�̂͑ȉ~�`�ŁA���a2�`3mm�̏��������̂��قƂ�ǁB�̂͌Q�̏�ʂ݂̂ɕ��z���A�ʏ�A�Q�̂̊���瓯�S�~��E�T���i�X�e���C�h�^�ɔz��B���������̔z��͒f���I�ŁA�̂̋��E���ŃR�������Ԗڏ�ɔ��B���邽�߁A���̔z�F�����Â炢���Ƃ�����B�u�ǁE�]��4���܂Ŕ��B���A���̌�����ˏo�x���͎����ɂ�炸�قړ����B1���E2���u�ǂɔ�ׂ�3���E4���u�ǂ͂킸���ɒZ���A�����ɒB���Ȃ��B�����͉~����`�ȉ~����B1���u�ǂ̓������Ǝ����Ƃ̊Ԃɕ����̃p�����`������邱�Ƃ�����B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�����H�ɏʒr���ʌ̊�A�ł������邱�Ƃ�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
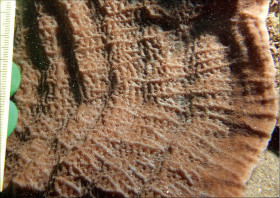
Leptoseris yabei (Pillai and Scheer, 1976)
�`�a�~�Z���x�C�T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Leptoseris Milne Edwards and Haime, 1849 �Z���x�C�T���S�� |
|

�B�e�F������^(����q���n���̊≮�A���[13m)
|

���ˏ�ɔ��B�����R�������悭�ڗ��B
|
|
�����`�F�t��`��Q�́B�Q�̂��قڕ��R�Ȃ��́A���S�~��ɔg�ł��́A�t����d�ɂ��d�Ȃ������̂�������B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Q�̎����ƃR�����㉏�͔����ۂ��Ȃ�B�����ȌQ�̂ł����S�̂͌����Ȃ��B
���i�̓����F�Q�̕\�ʂ̃R�����ɂ́A���ˏ�ɘA�Ȃ���̂Ɠ��S�~��ɘA�Ȃ���̂������A�R�������i�q��ɂȂ����Ƃ���̓�����1�`�����̌̂��T���i�X�e���C�h�^�ɔz��B���S�~��������ˏ�̃R�����̂ق����悭�ڗ��B�G���ȌQ�̂قǂ��̘A�������悭�A���S�~��ɔg�ł����Q�̂ł͘A�����������B�̂͑ȉ~�`�`���p�`�A���a2�`5mm�A�Q�̏�ʂ݂̂ɕ��z����B�u�ǁE�]��4���܂Ŕ��B����B1���E2���u�ǂ�3���E4���u�ǂ�������ɓˏo���A䰐S�߂��܂ł悭�L�т�B3���E4���u�ǂ�1���E2���ɔ�ׂĂ�┖���A䰐S�܂ŒB���邱�Ƃ͂Ȃ��B�����ׂ͍��~���A������B�Q�̗̂��ʂ͕�2.5mm�`1cm�̐���ɂȂ��Ă���A���������Ƃ���ł͐��̊Ԃ��[���ꍞ�݂ɂȂ�B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Pachyseris speciosa (Dana, 1846)
�����E�����T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pachyseris Milne Edwards and Haime, 1849 �����E�����T���S�� |
|

�B�e�F���{�@��(���V�\�s�匴�A���[10m)

|

���s����R�����̊Ԋu��2mm�O��B
|
|
�����`�F�t��`�핢��Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B
���i�̓����F�̂͌Q�̏�ʂ����ɕ��z�B�Q�̕\�ʂɂ́A�Q�̂̒��S���瓯�S�~��ɃR�������悭���B���A���s�ɕ��ԃR�����̊ԂɌ̂��z�邪�A�X�͕̌̂s���āB�R�����͌���1�`1.5mm�A����1�`2mm�قǁB���s����R�����̊Ԋu���Ȃ킿�̒��a��2mm�O��B�R�����̈ꕔ���r�ꂽ�Ƃ���ł́A�����e�B�N�������B���邱�Ƃ�����B�̂̊u�ǂ̓R�����������玲���܂Ŕ��B���A�قړ��������E�����Ŏ����̈Ⴂ�͕s���āA�R�����̖ʂɑ��Đ����ɓ��Ԋu�Ő��R�Ɣz��B䰐S�ɖ��ĂȎ������ώ@���邱�Ƃ͂ł��Ȃ����A�_��܂��͒Z����̎��������܂Ɍ����邱�Ƃ�����B�Q�̗��ʂ̎����ł́A�㉏�ɔ����ȋ��������]���悭���B����B�����̎����͕s�������A�����Ă��ˏo�����]�Ɣ����Ă��܂�ˏo���Ȃ��]�����݂ɕ��ԁB
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�ʒr���ʌ̊�A�ł������邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂��܂Ɍ�����B
�����EVeron (1995)�ł̏o���L�^�F���iP. minuta�Ƃ��ċL�^�j

|
|
| |
Pavona cactus (Forskål, 1775)
�T�I�g���V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F�쑺�b��(����q���剖���A���[5m)

|

�̂̒��a��1mm�قǂŏ������B
|
|
�����`�F�t��Q�́B�傫�ȗt�̐�[�����A����ɏ�����2�`3���̗t���`������B�܂��A�����̑������召�̗t���ꕔ�Z�����A��[���֍s�����傫�ȗt���`�����邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`�ΐF�B�t�̐�[�t�߂͑��̕������������ۂ��Ȃ�₷���B
���i�̓����F�傫�ȗt�̊�Ō���3�`3.5mm�A���⍂����5cm�قǂ܂Ő�������B���̐�[�t�߂ŁA�����܂��͓������ɕt�����鏬���ȗt�́A��Ō���2�`3mm�A���⍂����2cm�قǂ܂Ő�������B�̂͗t�̗��ʂɑ��݂��A�T���i�X�e���C�h�^�ɔz��B�傫�ȗt�ł́A�����̌̂��t�㉏�Ƃقڕ��s�ɘA�Ȃ��Ĕz��B�͉̂~�`�`�ȉ~�`�ŁA���a��1�`1.5mm�قǂŏ��������̂��������A�����̒��ɂ�1.5mm����傫�߂̌̂����݂��邱�Ƃ�����B�̂ɂ́A�Z���~����`�ȉ~����̎����ƁA1����2���̊u�ǁE�]���悭���B����B1���u�ǂ͎����܂ŒB����B2���u�ǂ�1���u�ǂɔ�ׂĔ����ĒZ���A�����ɒB���Ȃ��B�������A���a��1mm����傫�߂̌̂ł́A�����Z��3���u�ǁE�]�����B���A2���u�ǂ�1���u�ǂƂقړ��������ɂȂ��Ď����ɒB���邱�Ƃ�����B
�������F�ʒr�E��ʌ�A�g�Q�̉e�������܂�Ȃ��ʎΖʂ̐[�݂��ʎΖʂŌ�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Pavona decussata (Dana, 1846)
�V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|
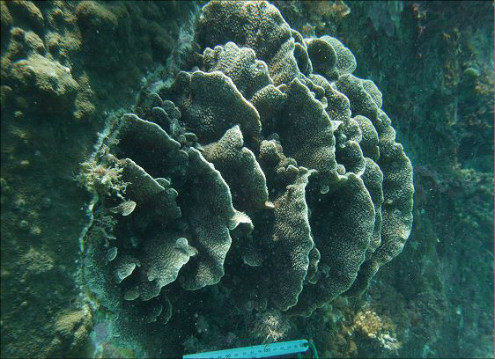
�B�e�F���{�@��(���V�\�s��v�ۍ`�A���[3m)

|
�̂̒��a��3�`5mm�Ƒ傫���B
|
|
�����`�F���߂̗t��`��Q�́B���������ɐL�т���`�̗t��`���A���ˏ�܂��͊i�q��ɔz�Ȃ��琬�����A���ꂢ�Ȕ�����̉��Q�̂��`�����邱�Ƃ������B�T���S�ʂ̏ʒr���ʌł́A�t���������핢��Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�B�W���F�`�����F�̌Q�̂�������B���ш�ł́A���Ԃł��G���L���Ă��邱�Ƃ������B
���i�̓����F�傫�ȗt�܂��͔̊�Ō���6mm�`1cm�قǁA���⍂��5�`10cm�قǁB�����̐�[�߂��Ő����܂��͓������ɕt�����鏬���ȗt�́A��Ō���3�`5mm�A���⍂��3�`5cm�قǂɐ�������B�̂͗t�܂��͔̗��ʂɑ��݂���B�����̌̂��Q�̎����Ƃقڕ��s�ɘA�Ȃ�Ȃ���T���i�X�e���C�h�^�ɔz��B�������A�傫���̈قȂ�̂����݂���Q�̂ł́A���̔z����邱�Ƃ������B�̂͒��a3�`5mm�قǂ̑ȉ~�`�����A�o��r���ŕ����̌����������̂������ꍇ�́A���傫��������B�̂ɂ�1�`3���̊u�ǁE�]���悭���B����B�����͊T���Ĕ��B�������A�܂�ɐj��܂��͔���̂��̂��`���������x�B1���E2���u�ǂ͂قړ��������ŁA䰐S�܂��͎����܂ŒB����B3���u�ǂ͔����ĒZ���A䰐S�܂ŒB���Ȃ��B
�������F�ʒr�E��ʌA���[15m�Ȑ�̒��ʂ��̂悢���p��g�Q�̉e���̏��Ȃ���ʎΖʂŌ�����B����{���Ȗk�ł́A�ʒr���ʎΖʂ̐��[5m�t�߂ő�Q�����`�����邱�Ƃ�����B
�����ł̕��z�F�É������ɓ��E���茧�ܓ����]���ȓ�B��q���ł͕��ʎ�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Pavona duerdeni Vaughan, 1907
�n�}�V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F����_�V(���V�\�s��Óc�A���[15m)
|

�̂̒��a��2�`3mm�Ə������B
|
|
�����`�F���_��`��Q�̂ŁA�Q�̎������t��ɂȂ邱�Ƃ͋ɂ߂ċH�B�召�����̞��_��`�����ɕ��сA����ɗאڂ��铯�l���Ƃقړ��Ԋu�œ������ɕ��ԁB���̂��߁A�S�̂Ƃ��Ă͔�����܂��͕s�K���ɐ���オ�������Q�̂��`�����邱�Ƃ������B
��̕��̐F�ʂƓ����F��ɊD���F�B�W���F�̌Q�̂�������B
���i�̓����F�͌���3�`5cm�A���⍂��10cm�ȏ�ɂȂ�B�����̗t��`���̂悤�ɁA���������ɐL�т�t���̔͂��܂�`������Ȃ��B�͉̂~�`�`�ȉ~�`�ŁA���a2�`3mm�A�T���i�X�e���C�h�^�ɔz�A�אڂ���̊Ԋu�͋����B�����Ȍ̂ł�2���u�ǁE�]�܂ŁA�傫�Ȍ̂ł�3���u�ǁE�]�܂Ŕ��B����B�������̊Ԋu���������߁A��������ɔ�ׂĘ]���Z���B�O�҂͞��_��`�̐�[�ő����A��҂͂����̑����ő���������B���ɑO�҂́A�אڂ���̓��m��䰕ǂ̈ꕔ�����L���ăZ���I�C�h�^�ɔz�邱�Ƃ������B�̂́A��[�̐�����~����`���G���ȑȉ~����̎��������B1���u�ǂ͌����A�����܂ŒB����B3���u�ǁE�]�����B�����̂ł́A2���u�ǂ�1���u�ǂƓ����x�̌����ɂȂ�A�قڑS�Ă������ɒB����B���̏ꍇ�A3���u�ǂ�2���u�ǂƂقړ������������A�����ɒB���邱�Ƃ͂Ȃ��B2���u�ǁE�]�܂ł������B���Ă��Ȃ��̂ł́A2���u�ǂ�1���u�ǂ����Z���A�����ɂ͒B���Ȃ��B
�������F�g������̋����ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō����A���ɏʎΖʂł́A���[5m�t�߂̉��r��Ō�����B
�����ł̕��z�F���m�������ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
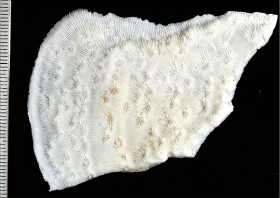
Pavona explanulata (Lamarck, 1816)
�q���V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F������^(���V�\�s��Óc�A���[11m)
|

�̂�䰕ǂ͕s���āB
|
|
�����`�F��ɔ핢��Q�̂ŁA�Q�̎����͗t��ɂȂ邱�Ƃ������B�܂��A�Q�̒��S���ɞ��_��̓ˏo�����s�K���Ɍ`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�`�Ԋ��F�B���Ԃł��G���L���Ă��邱�Ƃ������B
���i�̓����F�̂͒��a3�`5mm�̉~�`�`�ȉ~�`�ŁA�Q�̎����̂��̂قǑȉ~�`�ɂȂ�₷���B�o��r���̌̂͂��傫��������B�̔z��̓T���i�X�e���C�h�^�B�������A�Q�̒��S����ˏo���̒����ł́A�̊Ԋu�����ɂȂ��ăv���R�C�h�^�`�Z���I�C�h�^�z��ɂȂ邱�Ƃ�����B�Q�̎�����t�ł́A�Q�̂̒��S�ɋ߂��ق���䰕ǂ��킸���ɓˏo���A�̂��������������Ƃ������B�אڂ���̊Ԋu�͍L���A1cm�قNJJ�����Ƃ��������Ȃ��B�̂̊u�ǁE�]��3���܂Ŕ��B����B1���E2���u�ǂ͌����A�悭�ˏo���A�����܂ŒB����B3���u�ǂ͔������܂�ˏo�����A1���E2���u�ǂƋK�����������݂ɔz�A���̂قƂ�ǂ͎����܂ŒB���Ȃ��B�Q�̕\�ʂł́A1���E2���u�ǁE�]�̏㉏�͔����Ȃ��Ă���A����ł͂���炪����ɓˏo�����悤�Ɍ�����B�t�̗��ʂɌ̂͌`������Ȃ����A�����Ē���1���E2���]��3���]���悭���B���A����炪���݂ɔz��B�����͑傫���A�~�`�`�ȉ~�`�ŏ���ɂ悭�ˏo����B�o��r���̌̂ł́A��������ɒ����L�тĂ��邱�Ƃ������B
�������F�ʒr�E��ʌA���ʂ��̂悢���p�̐[�݂�g�Q�̉e���̏��Ȃ���ʎΖʂł��܂Ɍ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Pavona sp. �R�u�V�R���T���S�ƍ�������Ă���\��������A�����ł̐����ɂ��Ă͍Č������K�v�B
* ��q�����L�^��
|
|
| |
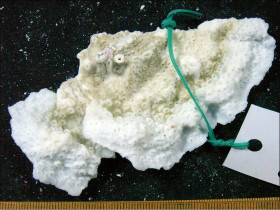
Pavona maldivensis (Gardiner, 1905)
�����W�u�V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F�[���T�L(����q���n���≮�A���[12m)

|

�̂��u��ɓˏo���₷���B
|
|
�����`�F��ɔ핢��Q�̂ŁA�Q�̎����͗t��ɂȂ邱�Ƃ������B�܂��A�Q�̒����ɏ����Ȟ���➞�_��̓ˏo���������̂��悭������B
��̕��̐F�ʂƓ����F�D�F�����������F��ΐF�B�t�̎����➶��`���_�̒����̐F�͒W���Ȃ�B���Ղ����F�Ȃǂ̈Ⴄ�F�ɂȂ����Q�̂������B
���i�̓����F�͎̂�ɒ��a2�`3mm�̉~�`�����A�Q�̎����ł͑ȉ~�`�ɂȂ邱�Ƃ�����B����`���_�̒����������A�אڂ���̊Ԃ͗���Ă���A�T���i�X�e���C�h�^�ɔz��B�������A�̂��u��ɓˏo���₷�����߁A�v���R�C�h�^�z��̂悤�Ɍ�����B�Q�̗̂��ʂɌ̂��`������邱�Ƃ͂Ȃ��B�Q�̏�ł̌̂̑傫����̊Ԋu�͕s�K���B�u�ǁE�]��3���܂Ŕ��B����B1���E2���u�ǂ͌������������قړ����ŁA�����܂ŒB����B3���u�ǂ͂���������┖���ĒZ���A�����ɂ͒B���Ȃ��B�����͉~����`�ȉ~����ŁA����`���_�̒����̌̂ł͖����B�̂��̂�����B
�������F�g������̎ア�ʎΖʂ̐��[20m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Pavona varians (Verrill, 1864)
�V���V�R���T���S |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F�쑺�b��(����q���n���≮�A���[6m)

|

�����I�Ƀ����e�B�N�����`�������B
|
|
�����`�F��ɔ핢��Q�̂ŁA�Q�̎������t��ɂȂ邱�Ƃ͂Ȃ��B��ՂɌŒ������A�C��̃T���S�I�Ȃǂ��Ȃ��琬���������Q�̂�������B
��̕��̐F�ʂƓ����F�����F�`���F��ΐF�B
���i�̓����F�̂͊p�����~�`�`���p�`�ŁA�T���i�X�e���C�h�^�ɔz��B�̂͒��a2�`5mm�A�召�l�X�Ȍ̂��Q�̏�ŕs�K���ɕ��z����B�אڂ���̊Ԃɂ́A�傫���̈قȂ郂���e�B�N����R�������s�K�����f���I�Ɍ`�������B�����̃R���������s�ɒ����A�Ȃ����Ƃ���ł́A�̂����A���h���C�h�^�z��̂悤�Ɍ�����B�u�ǁE�]�́A䰌a�̏����Ȍ̂ł�2���܂ŁA�傫�Ȍ̂ł�4���܂Ŕ��B�B䰌a�̏����Ȍ̂ł́A����1���u�ǂ݂̂������܂ŒB���A����2���u�ǂ͒B���Ȃ��B䰌a�̑傫�Ȍ̂ł́A2���u�ǂ�1���u�ǂƓ����x�̌����ɂȂ�A�Ƃ��Ɏ����܂ŒB����B3���E4���u�ǂ͂��ꂼ��2���E3���u�ǂ��������ĒZ���B�����͉~����`���G���ȑȉ~����B
�������F�ʒr�E��ʌ̐�A�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F���{�ȓ�B��q���ł͋H�B
�⑫�F�{���Pavona venosa �V�R���L�N���C�V�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Pavona sp.
�R�u�V�R���T���S�i�V�́j |
Agariciidae Lamarck, 1801 �q���t�L�T���S��
Pavona Lamarck, 1801 �V�R���T���S�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[5m)

|

�̂�䰕ǂ͖��āB
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂɒ��a�ƍ�������cm�قǂ̃R�u���s�K���Ɍ`�������B�܂��A�Q�̎������킸���ɗt��ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�ŁA�D�F�ɋ߂��F�ʂ̂��̂�����B���Ԃł��Q�̕\�ʂɂт�����ƐG�肪�L�тĂ���B
���i�̓����F�̂͒��a3�`5mm�̉~�`�`�ȉ~�`�ŁA�Q�̎����̂��̂قǑȉ~�`�ɂȂ�₷���B�̔z��͊�{�I�ɃZ���I�C�h�^�����A�Q�̎����ł͗אڂ���̂Ƃ̊Ԋu���L���Ȃ�A�T���i�X�e���C�h�^�ɂȂ邱�Ƃ�����B�u�ǁE�]��3���܂Ŕ��B�B1���E2���u�ǂ�䰕Ǐ㉏�ł悭�ˏo���A�����܂ŒB����B3���u�ǂ́A1���E2���u�ǂƔ�ׂ�Ɣ����A����1/3�`1/2�قǂŁA�����ɒB���邱�Ƃ͂Ȃ��B�Q�̗t�̗��ʂɌ̂͌`������Ȃ����A���������]�����B����B�����͎����ɂ��Ⴂ���s���ĂŁA�قƂ�Ǔˏo���Ȃ����߁A����ł͕�����ɂ����B�����͑傫���A�~�`�`�ȉ~�`�̖_��ŁA�o��r���̌̂ł́A�G���Ȕ�ɂȂ��Ă��邱�Ƃ������B
�������F�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�B��q���ł͋H�B
�⑫�F�{���Pavona explanulata �q���V�R���T���S��P. duerdeni �n�}�V�R���T���S�ƍ�������Ă���\��������A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
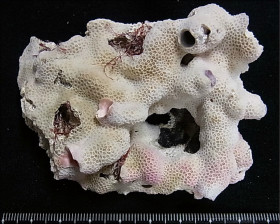
Stylocoeniella guentheri (Bassett-Smith, 1890)
���J�V�T���S |
Astrocoeniidae Koby, 1890 ���J�V�T���S��
Stylocoeniella Yabe and Sugiyama, 1935 ���J�V�T���S�� |
|

�B�e�F�o�H���q(���V�\�s��v�ۍ`�A���[9m)

|

䰌a��0.7�`1mm�B
|
|
�����`�F��ɔ핢��Q�̂ŁA�Q�̕\�ʂɒZ���}�����傫�Ȕ�����Q�̂��`�����邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�ŁA�|���v�̌��ՂƐG��̐�[�����F�Ȃǂ̔����F�ɂȂ�B�����ۂ�������̂��Q�̕\�ʂɖ��ɖ��ՂȂ����z����̂ŁA�Q�̕\�ʂ������ۂ��ڂ₯�Č�����B
���i�̓����F�̂͑��p�`�`�~�`�ŁA䰌a��0.7�`1mm�B�̊Ԋu�͂��s�K���ŁA1mm�ȏ�Ƃ��������A���ڂ���䰕ǂ̈ꕔ�����L����Ƃ��������B�u�ǂ�2���܂Ŕ��B���A䰕Ǐ㉏�ł悭�ˏo����B�u�Ǐ㉏�ɂ�3�`4�̋������A�u�ǂƋ����̑��ʂɂ͑�����������̑������`�������B1���u�ǂ�䰕Ǒ��ł͌������A䰐S���Ɍ������Ĕ����Ȃ�A�������k���ȉ~���`��M��̎����܂ŒB����B1���u�Ǔ����ɂ́A���������͂ނ悤��3�`6�̃p�����`�������B2���u�ǂ�1���u�ǂɔ�ׂĂ�┖���A�����ȉ��̒����Ŏ����ɒB���邱�Ƃ͂Ȃ��B�������k���Ō����A���̕\�ʂɂ͒Z�߂̒P���[�������B�B�܂��̂�䰕NJO���ɁA���a�܂��͍������ő��1mm�قǂ̔����ˋN���s�K���Ɍ`������邱�Ƃ�����B�}�����Q�̂̏ꍇ�A��1���}�̒�����cm�����Œ��a��1cm�O��A��2���}�̒�����1.5cm�����Œ��a8�`9mm�ɂȂ�B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʈ�A���p��ʒr�E��ʌŌ�����B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
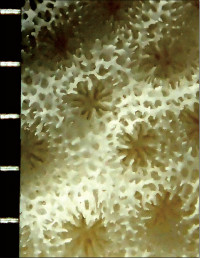
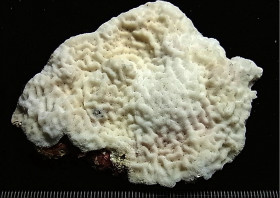
Coscinaraea columna (Dana, 1846)
���X���T���S |
Coscinaraeidae Benzoni, Arrigoni, Stefani and Stolarski, 2012 ���X���T���S��
Coscinaraea Milne Edwards and Haime, 1848 ���X���T���S�� |
|

�B�e�F�����@�O(���V�\�s��Óc�A���[10m)
|

䰂̒��a��2�`4mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̂̎�������Ղ���V�����āA����ɒ���o�����Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`�Ԋ��F�B�悭���B�����R�����̒��㕔�̗����ɁA���Ԃł��킸���ɐL�т��G��̐�[���A�����_����ɕ���Ō�����B
���i�̓����F�̂͑��p�`�ŁA䰂̒��a��2�`4mm�A�T���i�X�e���C�h�^�ɔz��B�אڂ���̊Ԃɂ͌�����2mm�قǂ̃R�������悭���B����B���̂���䰓����[���A�̂��Z���I�C�h�^�܂��̓��A���h���C�h�^�z��Ɍ�����B�u�ǁE�]��3���܂ŁA�����̏㉏�ɂ͐�[���������ɕ����������悭���B����B���̂��ߗאڂ���u�ǁE�]�Ԃ������A�Q�̕\�ʂ̓U���U�����Č�����B1�`3���u�ǁE�]�̌����ƍ����͂قړ����B1���E2���u�ǂ͒������قړ����ŁA�����Ȃ�Ȃ���䰐S�߂��܂ŒB����B3���u�ǂ̒�����1���E2���u�ǂ̔����ȉ��B䰐S�߂��ł́A1���u�ǂ̐�[����L�т��g���x�L���������ݍ����A���̏㕔�ɓݒ[�ȒZ���_��̎����ƕ����̒Z���p�����`�������B�������͎��͂̊u�Ǔ��������Ⴂ���߁A䰐S�ɂ͖��Ăȉ~�`�̒����|���`�������B�Q�̎����̗��ʂɂ̓G�s�e�J���`�������B
�������F�g������̎ア�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Coscinaraea monile (Forskål, 1775)
�m�}���X���T���S |
Coscinaraeidae Benzoni, Arrigoni, Stefani and Stolarski, 2012 ���X���T���S��
Coscinaraea Milne Edwards and Haime, 1848 ���X���T���S�� |
|

�B�e�F���{�@��(���V�\�s�Y�c�p�A���[6m)

|

䰂̒��a��4�`6mm
|
|
�����`�F�핢��`���Q�́B�Q�̂̎�������Ղ���V�����āA���t��`��ɒ���o�����Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`�Ê��F�B��u�ǁE�]�̏㉏��R�������㕔�������ۂ��A�U���U�����Č�����Q�̂������B���Ԃł��A��[�������A�����ĒZ����ׂ�̐G����o�����Ƃ�����B
���i�̓����F�̂͑��p�`�ŁA䰂̒��a��4�`6mm�قǂ̂��̂������B�אڂ���̊Ԃɂ́A������3�`4mm�A�������ő��3mm�قǂ̃R�����܂��̓����e�B�N�������B����B���̂��ߌ̔z��̓T���i�X�e���C�h�^�����A���ۂ̓Z���I�C�h�^�܂��̓��A���h���C�h�^�z��Ɍ�����B�u�ǁE�]��3���܂Ŕ��B���A�����͂ǂ���قړ����B�u�ǁE�]�̏㉏�ɂ́A��[���˂���A�������ɕ����傫�ȋ������悭���B����B����ČQ�̕\�ʂ̓U���U�����Č�����B�אڂ���u�ǁE�]�͂悭����A䰓��͐[��������B1���E2���u�ǂ́A�Ƃ��ɔ����Ȃ�Ȃ���䰐S�߂��܂ŒB����B䰐S�߂��ł́A1���u�ǂ̐�[����L�т��g���x�L���������ݍ����A�������`������B�������A�����̃g���x�L�����㕔�ɖ_��̎�����p�������B���邱�Ƃ͋H�B�����͎��͂̊u�Ǔ��������Ⴂ���߁A䰐S�ɂ͖��Ăȉ~�`�̒����|���`�������B3���u�ǂ͔��B���邪�A�Z����䰐S�߂��܂ł͒B���Ȃ��B�Q�̎����̗��ʂɃG�s�e�J�͌���ꂸ�A���ˏ�ɔ��B�����ׂ��]��������B
�������F�����������D�݁A�ʎΖʂł͐��[20m�Ȑ[�Ō�����B��ʎΖʂł͐��[10m�Ȑ[�A��ʈ�̓��p�ł͐��[10m�Ȑ�ł�������B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B
�⑫�F�{���Coscinaraea crassa �i�{�K�C�h���f�ڎ�j��Cycloseris explanulata �A�~���}���W���E�C�V�ƍ�������Ă���B����č����ł̐����ɂ��čČ������K�v�ł���B�{��̘a���Ƃ��̗R���͖쑺�E����(2005)���Q�ƁB
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Turbinaria frondens (Dana, 1846)
�E�l���X���o�`�T���S |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|

�B�e�F�k��T�q(���V�\�s��v�ۍ`�A���[9m)
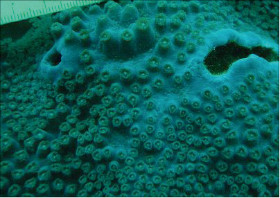
|

�u�ǐ���13�`18���قǁB
|
|
�����`�F�핢��`��Q�́B�Q�̂̒��a��50cm�ȏ�ŁA���S�t�߂̌�����2cm����Q�̂́A�Q�̎������傫���g�ł��āA�M�^�܂��͂��蔫�^�ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F�W���F�`���F�B���������Q�̂�������B
���i�̓����F�͉̂~�`�`���ȉ~�`�ŁA䰂̒��a��1�`1.5mm�B䰕ǂ͂悭�ˏo����B�핢��ɋ߂��Q�̂ł́A�̂�1.5�`5mm�قǕs�K���ɓˏo���A�~���`���ׂ�̉��ˌ^�Ɍ����邱�Ƃ�����B�̂̕��z�͋ψ�B���ꂢ�ȎM��`���蔫��Q�̂ł͌̂����S�~��ɔz�₷���B�אڂ���̊Ԃ̋����͋����B�u�ǂ�13�`18���قǂœ��Ԋu�ɔz�A䰐S�Ɍ������ċ}�X�ŗ������ށB�����́A�����̃p�����W�܂�����܂��͏����ȑȉ~�u�`�B�����͑��E�������A�E�̌`���Ԋu�͕s�����B�����\�ʂɂ͒P���[����ו���[�������B���邪�A�����̑傫����Ԋu�͕s�K���ŁA����炪����ɔz�邱�Ƃ͂Ȃ��B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂŌ�����B���܂ɐ�ʌΓ��Ō����邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B
�⑫�F�{���Turbinaria mesenterina �X���o�`�T���S�ƍ�������Ă���A���Ɏ�q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Turbinaria mesenterina (Lamarck, 1816)
�X���o�`�T���S |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|

�B�e�F�[���T�L(����q���剖���A���[2m)

|

�u�ǐ��͒ʏ�18�`20���قǁB
|
|
�����`�F�핢��܂��͗t��Q�́B���������Q�̎������s�K���ɏ������g�ł��Ƃ������A�Q�̒��S���ɉ~����̓ˋN�����`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`���F�B
���i�̓����F�͉̂~�`�`�ȉ~�`�A䰂̒��a�͒ʏ�1�`1.5mm�قǂ����A����^�̌̂��������Ă��邱�Ƃ�����B䰕ǂ��قƂ�Ǔˏo���Ȃ����߁A�̂͋����ɖ��݂��邩�킸���ɐ���オ����x�B�Q�̏�ł̌̂̕��z�́A�핢�ł͖��ŕs�K���A�t�ł͌�1�����x����Ăقڋψ�Ɍ�����B�����́A�����̃p�������ɕ���ŗZ��������܂��͑ȉ~�u�`�~�u�`�B�u�ǂ͒ʏ�18�`20���قǂ����A��^�̂ł�26�`28���قǂɂȂ邱�Ƃ�����B�u�ǂ�䰐S�Ɍ������ċ}�X�ŗ������ނ��A�������悭���B���Đ���オ��̂ŁA䰓��͂��قǐ[�������Ȃ��B䰕ǂ͑��E�������A�E���������ڗ����Ȃ��̂��k���Ȉ�ۂ���B䰍��\�ʂɂ͍ו���[���⑤�Ξ������B���A����ɔz��B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ�A��ʌΓ���ʒr���Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B
�⑫�F�{���Turbinaria frondens �E�l���X���o�`�T���S�AT. stellulata �q���X���o�`�T���S��T. sp. �C�{�X���o�`�T���S�Ȃǂƍ�������Ă���A���Ɏ�q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Turbinaria peltata (Esper, 1794)
�I�I�X���o�`�T���S |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|

�B�e�F�����C��(���V�\�s�Y�c�p�A���[6m)
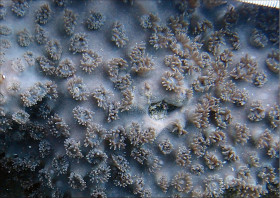
|
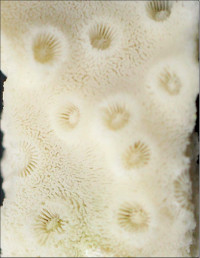
䰂̒��a��5�`6mm�Ƒ傫���B
|
|
�����`�F��ɔ�Q�̂ŁA���a50cm�A���S�t�߂̌���2cm����Q�̂́A�M�^�܂��͂��蔫�^�ɂȂ邱�Ƃ������B�܂��A�Q�̒��S���ɒ��a��3cm�O��̒������B���邱�Ƃ�����B����ɑ傫����������ƁA�Q�̂��o���̉ԕق̂悤�Ɋ�d�ɂ��d�Ȃ�A����Ȕ�����Q�̂ɐ������邱�Ƃ�����B
��̕��̐F�ʂƓ����F�D���F�`���F�B���Ԃł��G����悭�L���Ă���B
���i�̓����F�͉̂~�`�`���ȉ~�`�ŁA䰂̒��a��5�`6mm�Ƒ傫���B䰕ǂ͌����A�����͏����2�`5mm�قǓˏo���邽�߁A�͉̂~����ɓˏo���Č�����B�u�ǂ͒ʏ�24�������A�����24���̔��ɒZ���u�ǂ������̊Ԃɔ��B���邱�Ƃ�����B�u�ǂ̏㉏�Ƒ��ʂɂ́A�����ȋ�����������ˋN�����ꂼ�ꔭ�B����B�����͑傫�ȑȉ~�u�`�ŁA�����̊u�Ǔ������Ɍ`�����ꂽ�p����t�������ݍ������ƂŌ`�������B�����͑��E���ŁA�E�̌`���Ԋu�͕s�����B�����\�ʂɂ͒P���[����ו���[�����悭���B����B�܂��A�אڂ��邻���̞��̉����������I�ɗZ�����āA�����\�ʂ̈ꕔ������Ɍ����邱�Ƃ�����B
�������F�ʎΖʂ̐[���A����������ʈ��Օ��I�ȓ��p���Ō�����B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B
* ��q�����L�^��

|
|
| |
Turbinaria reniformis Bernard, 1896
���R�~�]�X���o�`�T���S |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|

�B�e�F�k��T�q(���V�\�s��v�ۍ`�A���[4m)

|

�u�ǐ���16�`20���ŒZ���B
|
|
�����`�F�핢��A�t��`��Q�́B�����`���ߏ�����ɐL�т������̗t�܂��͔��d�Ȃ荇�����Q�̂�����B
��̕��̐F�ʂƓ����F��ɉ����F�����A䰓��ƌQ�̎����݂̂����F���Ȃ������F�̌Q�̂�����B
���i�̓����F�̂͂قډ~�`�ŁA䰂̒��a��1.5�`1.8mm�A䰕ǂ��̂����܂�ˏo���Ȃ��B�̂͌Q�̕\�ʂɂقڋϓ��ɕ��z���A�אڂ���̂̊Ԋu�͌�1��������ȏ㗣��Ă��邱�Ƃ������B�u�ǂ�16�`20���ŒZ���A䰐S�Ɍ������Ăقڐ����ɗ������ށB���̂���䰓��͐[���A���������L��������B�����͕����̃p������`������A���̕\�ʂ͕��R���A���~�u��ɐ���オ��B�����͑��E���ŁA�~�`�̍E���قړ��Ԋu�ŊJ���Ă���B�����\�ʂ̞��͎�ɍו���[���ŁA�����I�ɒP���[�������݂��邱�Ƃ�����B�܂��A���������I�ɗZ�����Đ���Ɍ����邱�Ƃ�����B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʈ�̐��ʌŎ�Ɍ�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B
�⑫�F�{���Turbinaria frondens �E�l���X���o�`�T���S�ATurbinaria sp.�AT. mesenterina �X���o�`�T���S��T. stellulata �q���X���o�`�T���S�Ȃǂƍ�������Ă���A���Ɏ�q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FEuphyllia ancora�Ƃ��ċL�^�����\������

|
|
| |
Turbinaria stellulata (Lamarck, 1816)
�q���X���o�`�T���S |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|
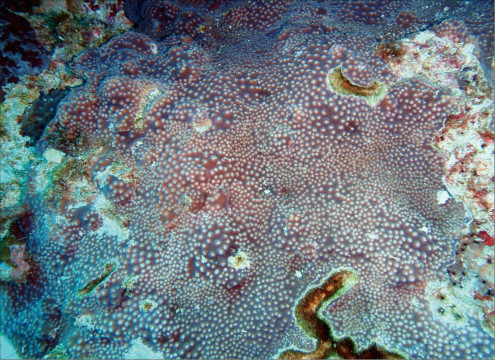
�B�e�F�쑺�b��(���V�\�s��Óc�A���[10m)

|

�u�ǐ��͒ʏ�24���O��B
|
|
�����`�F�핢��܂��͏����Q�́B�Q�̂̈ꕔ���R�u��ɐ���オ������A��������ɒ���o�����肷�邱�Ƃ�����B�Q�̂̒��S�߂��ł̌�����2cm�ȏ�ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�Ê��F�B䰓��܂��͌Q�̑S�̂������F�ɂȂ����Q�̂�������B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA䰂̒��a��1.5�`1.8mm�قǁB䰕ǂ�2mm���x�܂œˏo���邪�A䰕NJO�����悭������邽�߁A�̑S�͉̂~���`�Ƃ������h�[���^�ɐ���オ���Č�����B�Q�̕\�ʂ��R�u��ɐ���オ���������ł́A�̂͑傫���A�悭�ˏo���A�אڂ���̊Ԋu���L���Ȃ�B����A�Q�̎����ł́A�̂͏������A���܂�ˏo�����A�אڂ���̊Ԋu�������Ȃ�B�u�ǂ�18�`30���قǂŁA�ʏ��24���O�オ�����B�����u�ǂ̒��ɂ́A�Z���āA�אڂ����u�Ǒ��ɘp�Ȃ��Ȃ���Z��������̂�������B���̂��ߊu�ǂ̒����Ɣz���s�����Ȉ�ۂ���B䰕ǂ��悭�ˏo���Ă���A�u�ǂ�䰐S�Ɍ������ċ}�X�ŗ������ނ̂ŁA䰓��͐[��������B�����͕����̃p������`������邪�A���B�̓x���͗l�X�B�悭���B���������͑ȉ~�u�`�`�~�u�`�ɂȂ�B�����͑��E�������A�E�͏������k���Ɍ�����B�����\�ʂɂ͍ו���[�������W���A�����I�ɒP���[�������݂���B�܂��A���������I�ɗZ�����A����Ɍ����邱�Ƃ�����B
�������F��Ɋ�ʈ���ʌ̐Ō�����B
�����ł̕��z�F��q���ȓ�B
�⑫�F���F�̌Q�̂�Turbinaria frondens �E�l���X���o�`�T���S�ƁA�����F�̌Q�̂�Turbinaria reniformis ���R�~�]�X���o�`�T���S�ƍ�������Ă���A���Ɏ�q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Turbinaria sp.
�C�{�X���o�`�T���S�i�V�́j |
Dendrophylliidae Gray, 1847 �L�T���S��
Turbinaria Oken, 1815 �X���o�`�T���S�� |
|

�B�e�F�[���T�L(����q���n���̊≮�A���[6m)

|

�u�ǐ���13���O��Ə��Ȃ��B
|
|
�����`�F�핢��܂��͗t��Q�́B�Q�̕\�ʂ͕s�K���ɔg�ł��Ƃ������B
��̕��̐F�ʂƓ����F�W���F�`���F�B
���i�̓����F�͉̂~�`�ŁA䰂̒��a��1�`1.3mm�B䰕ǂ��ʏ�1�`1.5mm�قǓˏo���A�͉̂����Ԃ��ꂽ�悤�ȉ~���`�Ɍ�����B�܂��5mm�߂��ˏo�����̂�������B�Q�̕\�ʂł̌̂̕��z�͕s�K���ŁA�̓��m�����ڂ��Ȃ��瓯�S�~��ɔz�邱�Ƃ�����A5mm�O�㗣��Ă��邱�Ƃ�����B�u�ǂ�13���O��Ə��Ȃ��A���Ԋu�ɔz��B������䰕Ǒ��ł������A䰐S���Ɍ������Ĕ����Ȃ�Ȃ���}�X�ŗ������ށB�����͕����̃p�����W�܂��Ăł����~�u�`�ŁA���������̂̓���ł��悭�F���ł���B�������A�p���̔��B�������A�_��ɂ��������Ȃ��̂�������B�����͑��E���ŁA�����ȉ~�`�̍E�����Ԋu�ŋK���������J���Ă���A�X�|���W��Ɍ�����B�����\�ʂɂ͒P���[����ו���[�����悭���B����B���̑傫����Ԋu�͂悭�����B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʈ�̐��ʌŎ�Ɍ�����B
�����ł̕��z�F�����_�ł͎�q���݂̂���m���Ă���B�������{���Turbinaria frondens�E�l���X���o�`�T���S�AT. mesenterina�X���o�`�T���S��T. reniformis���R�~�]�X���o�`�T���S�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Euphyllia fimbriata (Spengler, 1799)
�i�K���n�i�T���S |
Euphylliidae Alloiteau, 1952 �n�i�T���S��
Euphyllia Dana, 1846 �i�K���n�i�T���S�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[12m)

|
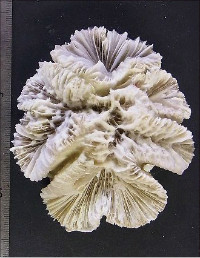
�t���x�����A���h���C�h�^�̌Q�́B
|
|
�����`�F�ԑ���Q�́B�������猩��Ɣ����`�̉��Ɍ�����B���a1m���邱�Ƃ��������Ȃ��B
��̕��̐F�ʂƓ����F��ɗΐF�ŁA�D�F�܂��͉��F�����������邢�F�ɂȂ�B�G��͒��Ԃł��悭�L������B�G���[�͐t���^(�܂��͕d�^)�ŁA�ʏ�ɂȂ������͔����ۂ��Ȃ邱�Ƃ������B���}�̑��ʂ́A�����܂œ����F�̋����ŕ����邱�Ƃ������B
���i�̓����F�̔z��́A�t�@�Z���C�h�`�t���x�����A���h���C�h�^�B���}��[�̌̂́A�O�������O���ɊJ�����~�`�`�ȉ~�`�ŁA���䰂̒��a��2�`3cm�B䰐S�Ɏ������������A����������������䰓���4�ȏ㑶�݂���䰂����тɂȂ�A���a��5cm���邱�Ƃ��悭����B5���u�ǁE�]�܂Ŕ��B���A�������Ƃɍ����ƒ������قȂ�B�����̏㉏�Ƒ��ʂɂ͋�����������̑����͂قƂ�ǔ��B���Ȃ��B1���u�ǂ̏㉏��䰕Ǐ㉏�ŏ���ɂ悭�ˏo���A��������䰐S�Ɍ������ċ}�X�ŗ������ށB�܂��A䰐S�߂��ł�1���u�ǂ̓��������p�Ȃ���B2���u�ǂ̌`������l�����A1���u�ǂقǂ͓ˏo�����A䰐S�܂ŒB���Ȃ����̂�������p�Ȃ��Ȃ����̂�������B�]�����l��5���܂Ŕ��B���邪�A5���]�������B�̌̂�����B1���E2���u�ǂ͒���1cm���A�f���I�ɌQ�̂̊�܂ŐL�т邱�Ƃ����邪�A����ȊO�̘]�͒���1cm�ɖ����Ȃ����̂��قƂ�ǁB
�������F���̑��������I����D�ށB�g�����肪�ア�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō����A���p��ʒr�E��ʌɂ��H�ɐ������邱�Ƃ�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʈȓ�B��q���ł͋H�B
�⑫�F�{���Euphyllia ancora �ƍ�������Ă���A�����ł̐����̍Č������K�v�ł���BE. ancora �͌̔z�t���x�����A���h���C�h�^�݂̂ŁAE. fimbriata �ɔ��䰂̒Z�a��������(�Q�̏�Ɍ`�����ꂽ�����J�̕�������)�A�u�ǐ������Ȃ��B�i�K���n�i�T���S�̘a����S���͖̂{��E. fimbriata �ł��邱�Ƃ���AE. ancora �̐V�̘a�����A�ׂ������J�������ƂɈ���Ńz�\�i�K���n�i�T���S�Ƃ���B
* ��q�����L�^��
|
|
| |
Euphyllia paraglabrescens Veron, 1990
�n�i�T���S���h�L |
Euphylliidae Alloiteau, 1952 �n�i�T���S��
Euphyllia Dana, 1846 �i�K���n�i�T���S�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

�̔z��͊�{�I�Ƀt�@�Z���C�h�^�B
|
|
�����`�F�ԑ���Q�́B�������猩��Ɣ����`�̉��Q�̂Ɍ�����B���a15cm�قǂ܂Ő�������B
��̕��̐F�ʂƓ����F��ɗΐF�ŁA�D�F�܂��͉��F�����������邢�F�ɂȂ�B�܂������Q�̓��ł������I�ɐF���قȂ邱�Ƃ�����B�G��͒��Ԃł��悭�L������B�G��̐�[�͋��`�ŁA���̗֊s�͔����ۂ��B���}�̑��ʂ́A�����܂œ����F�̋����ŕ����邱�Ƃ������B
���i�̓����F�̂̓t�@�Z���C�h�^�ɔz��B���}��[�̌̂́A�O��������̂悤�ɂ悭�J�����~�`�`�ȉ~�`�ŁA���䰂̒��a��2�`4cm�A�����������B�o��r���̏ꍇ�́A����������������䰓���2�`3���݂��邱�Ƃ�����B5���u�ǁE�]�܂Ŕ��B���A�������Ƃɍ����ƒ������قȂ�B�����̏㉏�Ƒ��ʂɂ͋�����������̑����͂قƂ�ǔ��B���Ȃ��B1���u�ǂ̏㉏��䰕Ǐ�[�ł悭�ˏo���A䰐S�߂��܂ł͊ɂ₩�ɌX������A䰐S�Ɍ������ċ}�X�ŗ������ށB�܂��A䰐S�߂��ł�1���u�ǂ̓��������p�Ȃ���B2���u�ǂ����l�̌`������Ă��邪�A1���u�ǂقǂ͓ˏo�����A䰐S�܂ŒB���Ȃ����̂�������p�Ȃ��Ȃ����̂�����������B�]�����l��5���܂Ŕ��B����B䰕ǂ̊O���ł̘]�̍�����1mm�قǂŁA1���E2���u�ǂ̒�����1cm���A�f���I�ɌQ�̂̊�܂ŐL�т邱�Ƃ����邪�A����ȊO�̘]�͒���5mm�ɖ����Ȃ����̂��قƂ�ǁB
�������F���̑��������I����D�ށB�g�����肪�ア�ʎΖʁE��ʎΖʂ�A���p�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���̂݁B��q���ł͋Ǐ��I�ɕ��ʎ�B
�⑫�F�{��̃^�C�v�Y�n�͎�q���剖���B
*** ����܂Ŏ�q���Ŋm�F����Ă��Ȃ���
|
|
| |
Galaxea fascicularis (Linnaeus, 1767)
�A�U�~�T���S |
Euphylliidae Alloiteau, 1952 �n�i�T���S��
Galaxea Oken, 1815 �A�U�~�T���S�� |
|

�B�e�F����_�V(����q���n���̊≮�A���[20m)

�u�ǂ�䰕Ǐ㉏�ʼns���ˏo����B |

|
|
�����`�F�핢��`���Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���ΐF�A���F�`�Ԋ��F�ȂǁB䰂̏���ɂ悭�L�т��u�ǂɂ���āA�X�̌̂��A�U�~�̉ԕُ�Ɍ�����B�G��͒��Ԃł��悭�L�сA���̐�[�͔���������B
���i�̓����F�̂̔z��̓v���R�C�h�^�B�͉̂~�`�`�p�����ȉ~�`�ŁA�Q�̂̒��S�t�߂̌̂قǑ傫���A�����̂��̂قǏ������B�Q�̒��S���ł̌̂́A���a��5�`7mm�A�A��̋����\�ʂ���8mm�`1cm�قlj~���`�ȉ~����ɂ悭�ˏo����B�אڂ���̂̊Ԋu��2�`3mm�B���̂悤�ɌQ�̏�ł̌̂̑傫���A�ˏo�x����Ԋu���悭�����̂ŁA�̂͐��R�Ɣz�Ă���悤�Ɍ�����B�u�ǂ�3���܂Ŕ��B���A�u�ǂ͂ǂ��䰕Ǐ㉏�Ō����A����ɉs���ˏo����B������䰕Ǐ㉏����䰐S�Ɍ����Ĕ����Ȃ�Ȃ���}�X�ŗ������ށB1���E2���u�ǂ�䰐S�܂ŒB���邪�A3���u�ǂ͂����̔����قǂ̒�����䰐S�ɂ͒B���Ȃ��B�ʏ�A�����͖����B�����A�̂ɂ���Ă�1���E2���u�ǂ̓��������p�Ȃ��Ȃ��������A�אڂ�����̂Ɩ������Ď�����̍\�����`��������̂�����B�]�͂킸���ɔ��B���邪�ڗ����Ȃ��B�Q�̗̂��ʂł́A���S�~��ɔ��B�����G�s�e�J��������B
�������F�T���S�ʂ̏ʒr�E��ʌ`�ʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Galaxea astreata �i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����̍Č������K�v�B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FDiaseris distorta�Ƃ��ċL�^�����\������
|
|
| |
Cycloseris costulata (Ortmann, 1889)
�X�W�}���W���E�C�V |
Fungiidae Dana, 1846 �N�T�r���C�V��
Cycloseris Milne Edwards and Haime, 1849 �}���W���E�C�V�� |
|

�B�e�F������^(���V�\�s��Óc�A���[16m)

|

���ʂɂ͘]���悭���B����B
|
|
�����`�F���n�͔̂�Œ����̒P�̂ŁA�͉̂~�Ռ^�`���ȉ~�Ռ^�ɂȂ�B������������オ������A�������킸���ɔg�ł����肷�邱�Ƃ�����B�̂̒��a�͒ʏ�5cm�O��̂��̂��������A10cm�߂��܂Ő������邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`���F�B�\�ʂɈÊ��F�܂��͔����ۂ����䂪�t�����Ƃ������B
���i�̓����F�̂̕\�ʂ́A�������璆�S�ɂ����ăh�[����܂��͂��~����ɐ���オ��A���S����1�`1.5cm�قǂ̍����ɂȂ�B���Ղ��܂ޒ����|�̒��a�́A�̂̒��a��1/6�قǂ܂ŁB�u�ǂ͒ʏ�6���܂Ŕ��B���邪�A��^�̌̂ł͕����I��7���u�ǂ܂Ō`������邱�Ƃ�����B�u�ǂ̒����͎������Ƃɖ��ĂɈقȂ�A�����̂��̂قǒZ���B�������A5���u�ǂ�1���E2���u�ǂƂقړ��������ɂȂ�A�����|�܂ŒB����B1�`4���u�ǂ͌����ƍ��������܂�ς�炸�A�����|���ł͔�����ă��[�u��ɓˏo������A���E�ɔg�ł����肷�邱�Ƃ�����B����A5���E6���u�ǂ͔����A�ˏo���Ȃ��B�̗̂��ʂɂ͘]���悭���B����B�����͎����Ɋւ�炸�قړ��������ŁA�قڂ܂������ɐL�тĂ���A���E�ɑ傫���g�ł��Ƃ͂Ȃ��B�]�̏㉏�ɂ́A��[�̐���������ȋ��������R�ƕ��ԁB�̕ǂ��k���ōE�͊J�����A�\�ʂ͕����Ɍ�����B
�������F�g������̎ア�ʎΖʂ��ʎΖʂ̍��I��Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Cycloseris tenuis �}���W���E�C�V���h�L��C. vaughani �i�{�K�C�h���f�ڎ�j�ƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FC. vaughani�Ƃ��ċL�^�����\������
|
|
| |
Cycloseris explanulata (van der Horst, 1922)
�A�~���}���W���E�C�V�i�V�́j |
Fungiidae Dana, 1846 �N�T�r���C�V��
Cycloseris Milne Edwards and Haime, 1849 �}���W���E�C�V�� |
|

�B�e�F�����@�O(���茧��ܓ����ʓ��A���[15m)

|

�̂̒��a��2.5�`4mm�ŕs�����B
|
|
�����`�F�Œ����̔핢��Q�́B�Q�̎����͊�Ղ��痣��邪�A�t��ɂȂ��ď���ɂ߂���邱�Ƃ͂Ȃ��B
��̕��̐F�ʂƓ����F���ΐF�`�ΐF�B�����ł��悭���W�E�ˏo����1���u�ǂ�F���ł���B���Ԃł���[�̔����G����킸���ɐL���Ă��邱�Ƃ�����B
���i�̓����F�̂͒��a2.5�`4mm�̊p�����~�`�`�ȉ~�`�ŁA�����̔z��̓T���i�X�e���C�h�^�ɂȂ�B�Q�̏�ł̌̂̑傫����Ԋu�͕s�����B�u�ǁE�]��3���܂Ŕ��B���A��̂��̂قnj����A䰕Ǐ㉏�t�߂ł悭�ˏo����B�܂������̏㉏�ɂ͕����̐�[�����������A���ʂɂ͐�[�̐���������ˋN���悭���B����B���̂��߁A����ł͌Q�̕\�ʂ��U�����Č�����B1���E2���u�ǂ͎����܂ŒB����B3���u�ǂ͎����܂ŒB�����A���̎�O��2���u�ǂɗZ������B1���u�ǂ̒��ɂ́A�אڂ���̂�1���]�łȂ��炸�ɁA�u�Ǐ㉏�Ō���������ĉԕُ�ɂȂ邱�Ƃ�����B�����́A�u�Ǔ�������L�т������̃p���Ɩ_��ˋN�Ō`������邪�A�p���������B�̌̂������B�̊Ԋu���L���Ƃ���ł́A�אڂ���]�Ԃ��i�q��ɂȂ��V�i�v�e�B�L�������悭���B����B���̂��ߋ����\�ʂ����ꂢ�ȖԖڏ�Ɍ�����B�]�́A�Q�̗̂��ʂł��m�F�ł��邪�A�����͕s���Ăłǂ���ڗ����Ȃ��B
�������F��ʎΖʂ̐��[15m�Ȑ[�A�܂��͓��p�̐��[5m�Ȑ[�̊�Տ�⋐�I��Ō�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʓ��ȓ�B��q���ł͋L�^����Ă��Ȃ��B
�⑫�F�{��̐V�̘a���́A�̔z�T���i�X�e���C�h�^�ŁA�אڂ���u�NJԂ������̃V�i�v�e�B�L�����Ŋi�q��ɘA�Ȃ��ĖԖڏ�Ɍ����邱�ƂɗR������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Cycloseris sinensis Milne Edwards and Haime, 1851
�V�i�}���W���E�C�V |
Fungiidae Dana, 1846 �N�T�r���C�V��
Cycloseris Milne Edwards and Haime, 1849 �}���W���E�C�V�� |
|

�B�e�F��������(�B�e�n�_�Ƃ��̐��[�A�W�{�̏��݂͕s��)

�������ʐ^�̌Q�̂Ƃ͕ʌQ��(����������E���u�ˉ��A���[30m�ō̏W)�B |

|
|
�����`�F���n�͔̂�Œ����̒P�̂ŁA�͉̂~�Ռ^�̂��̂����邪�A�ǂ��炩�Ƃ����ƕ��ˏ�̐ꍞ�݂��������ԕٌ`��A����炪����Đ�`��t���`�ɂȂ������̂������B�̂̒��a�͑傫�Ȃ��̂�3cm�O��B
��̕��̐F�ʂƓ����F�W���F�`���F�B�����u�Ǐ�ɈÊ��F�܂��͔����ۂ������Ȕ��䂪�t���B
���i�̓����F�̂͝G�����A���S������オ��A���S���̍���1cm�����̂��̂��قƂ�ǁB�����|�̒��a�́A�̂̒��a��1/5�قǂ܂ŁB�u�ǂ͒ʏ�7���܂Ŕ��B����B�����̏㉏�Ƒ��ʂɂ́A������̑������t���������Ɣ����ˋN���悭���B���A���R�Ɩ��ɔz��B�u�ǂ̒����͎������Ƃɖ��ĂɈقȂ邪�A�����⍂���͂V���u�ǂ������قƂ�Ǖς��Ȃ��B1�`6���u�ǂ͌����A���ɓ����Ɍ������قǂ悭�������B�܂������́A�ɂ₩�ɔg�ł��Ă��邱�Ƃ������B7���u�ǂ͔����A�אڂ���1�`6���u�ǂ̊Ԃ��[�U����悤�ɔ��B����B���̊u�ǂ́A3�`6���u�ǂƂ����̓����ŗZ�����Ă���B7���u�ǂ̋����͑��̊u�ǂ̂����ɔ�ׂđ傫�����߁A7���u�ǂ̏㉏�͓_����ɂ݂���B�̗̂��ʂł͕\�ʂ̊u�ǂɑΉ������]���悭���B����B�����������͓ˏo������g�ł����肷�邱�Ƃ͂Ȃ��A���S�ɂ����قǔz�s���ĂɂȂ�B�]�̏㉏�ɂ͐�[�̐���������ȋ������K���������z��B�̕ǂ��k���ōE�͊J���Ȃ��B
�������F�ʎΖʂ��ʎΖʂ̐[�݂̍��I��Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Cycloseris distorta �i�{�K�C�h���f�ڎ�j�ƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F���iFungia scutaria�Ƃ��ċL�^�j
|
|
| |
Cycloseris tenuis Dana, 1846
�}���W���E�C�V���h�L |
Fungiidae Dana, 1846 �N�T�r���C�V��
Cycloseris Milne Edwards and Haime, 1849 �}���W���E�C�V�� |
|

�B�e�F������^(���V�\�s��Óc�A���[17m)

|

���ʂł͒�u�ǂ̂ق����ڗ��B
|
|
�����`�F���n�͔̂�Œ����̒P�̂ŁA�͉̂~�Ռ^�`���ȉ~�Ռ^�ɂȂ�B������������オ������A�������킸���ɔg�ł����肷�邱�Ƃ�����B�̂̒��a�͒ʏ�5cm�O��̂��̂��������A10cm�߂��܂Ő������邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`���F�B�̕\�ʂɈÊ��F�܂��͔����ۂ����䂪�t�����Ƃ������B
���i�̓����F�̂̕\�ʂ́A�������璆�S�ɂ����ăh�[����܂��͂��~����ɐ���オ��A���S����1�`2cm�قǂ̍����ɂȂ�B�����|�̒��a�́A�̂̒��a��1/5�قǂ܂ŁB�u�ǂ͒ʏ�6���܂Ŕ��B���邪�A��^�̂ł�7���u�ǂ܂Ō`�������B�u�ǂ̒����͎������Ƃɖ��ĂɈقȂ�A�����̂��̂قǒZ���B������6���u�ǂ́A�܂�5���u�ǂ̓����ŗZ�����A����4���u�ǂ̓����ŗZ�����A�����3���u�ǂ̓����ŗZ�����Ȃ���1���E2���u�ǂƂقړ��������ɂȂ�A�����|�܂ŒB����B1�`5���u�ǂ͌����ƍ��������܂�ς�炸�A�����|���Ŕ�������胍�[�u��ɓˏo������A�킸���ɍ��E�ɔg�ł����肷�邱�Ƃ�����B����A6���u�ǂ͔����ˏo���Ȃ��B�̗̂��ʂɂ́A�܂������A�܂��͂��g�ł����]���悭���B����B�]�͒��S�Ɍ������ɂ�ĕs���ĂɂȂ�B�]�㉏�ɂ́A��[����葤�ʂ�������̑������t���������ȋ��������s�K���ɕ��ԁB������1�`4���]�Ō����ɔ��B���邽�߁A1�`4���]��5���E6���]�ɔ�ׂ�Ƃ悭�ڗ��B�̕ǂ��k���ōE�͊J�����A�\�ʂ͕����Ɍ�����B
�������F�g������̎ア�ʎΖʂ��ʎΖʂ̍��I��Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�}���W���E�C�V���h�L�̘a����S���͖̂{��B�{��͂���܂�Cycloseris costulata �X�W�}���W���E�C�V��C. vaughani �i�{�K�C�h���f�ڎ�j�ƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Lithophyllon undulatum Rehberg, 1892
�J�����T���S |
Fungiidae Dana, 1846 �N�T�r���C�V��
Lithophyllon Rehberg, 1892 �J�����T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[6m)

|

�����̂�䰂̒��a��4mm�`1cm�B
|
|
�����`�F�T���S�ʈ�ł͂��тȑȉ~�`�̔핢��Q�̂ɁA���ш�ł͐��\cm����傫�ȗt��܂��͔�Q�̂ɂȂ�B�t��E��Q�̂͊�d�ɂ��d�Ȃ�Ȃ��琬������B�����̌Q�̂́A�[��ł͐��������ɁA���ł͌Q�̎������g�ł��A�ߏ�����ɐ������邱�Ƃ������B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�B���ΐF�̖��邢�Q�̂�������B���Ԃł��~����̑����G���5mm���x�L���Ă��邱�Ƃ�����B
���i�̓����F�̂͑ȉ~�`�ŁA�Q�̏�ʂ݂̂ɕ��z����B�c�Q�̂ł͒��S�̂�F���ł��邪�A�Q�̂̐����ƂƂ��Ɏ����̂����F���ł��Ȃ��Ȃ�B�����̂�䰂͒��a��4mm�`1cm�A�̂̐���傫���A�Ԋu�͌Q�̂ɂ���ĈقȂ�B�����̂̓T���i�X�e���C�h�^�ɔz�A�����I�ɓ��S�~��ɔz��B�����̂̊u�ǁE�]��4���܂Ŕ��B���A�㉏�ɂ͞���̑��������ג����������悭���B����B1���u�ǁE�]�ł́A�����̋����Ԃ������I�ɗZ������B1����2���̊u�ǁE�]�͌����A�悭�ˏo���邪�A3����4���̊u�ǁE�]�͔����Ă��܂�ˏo���Ȃ��B1���E2���u�ǂ̓����́A����Ǝ������Ɍ������ă��[�u��ɒ���o���Ȃ��玲���ɒB����B3���u�ǂ͎����܂ŁA4���u�ǂ͎�����O�܂Ŕ��B����B�܂�4���u�ǂ́A������O��3���u�Ǒ��ɘp�Ȃ��Ȃ���Z������B�����͑傫���Ė��āB�����́A���������ɐL�т���̘]�Ƃ����𐅕��ɘA������_��̃V�i�v�e�B�L��������Ȃ�B���̃V�i�v�e�B�L�����́A�]�̉����œ��Ԋu�ɋK���������z��B�Q�̗��ʂɂ͘]�����B���邪�A�����̏㉏�ɋ����͂��܂蔭�B���Ȃ��B�̕ǂ��k���ŕ\�ʂ͕����Ɍ�����B�̕ǂɍE���J�����Ƃ͂Ȃ��B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[10m�Ȑ[�A���l�̊�ʎΖʂł͐��[5m�Ȑ[�Ō�����B���p�̑��������ł́A���[5m�Ȑ�ł�������B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
* ��q�����L�^��
|
|
| |
Lobactis scutaria (Lamarck, 1801)
�N�T�r���C�V |
Fungiidae Dana, 1846 �N�T�r���C�V��
Lobactis Verrill, 1864 �N�T�r���C�V�� |
|

�B�e�F�i�c�r��(���V�\�s���Y�A���[10m)
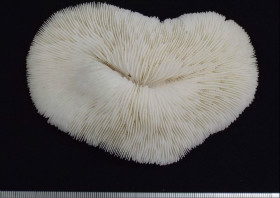
|

�����|�͑傫���B
|
|
�����`�F���n�͔̂�Œ����̒P�̂ŁA�̂͒��������h�[����ɐ���オ�������тȑȉ~�Ռ^�`�唻�^�ɂȂ�B�ʏ�͒��a��15cm�����̌̂��������A����ȏ�ɐ���������̂�����B
��̕��̐F�ʂƓ����F��Ɋ��F�`�W���F�B�G�肪�L�����镔���͎�����������ۂ��Ȃ��Ă���A�����ł��悭�ڗ��B���Ԃł��A��ׂ�̐G�肪��������킸���ɐL�тĂ��邱�Ƃ�����B
���i�̓����F�̕\�ʂ́A�������璆�S�ɂ����ăh�[����ɐ���オ��A���S����3�`5cm�قǂ̍����ɂȂ�B�̂̎����͂�◠�ʑ��Ɋ������܂��B�������̎����ɂ͕����̐ꍞ�݂��s�K���ɓ��邽�߁A���̗֊s�͔g�ł��Č�����B�����|�̒��a�͌̂̒��a��1/3�`2/5�قǁB�����͗l�X�ȕ����������������̃g���x�L��������Ȃ�B�u�ǂ͎����Ɋւ�炸�قړ��������ŁA�����|�Ɍ������Ĕg�ł��Ȃ�����ˏ�ɔz��B�u�Ǔ����́A��u�ǂقǒ����|���ɓˏo���邪�A�u�NJO���ł͎����ɂ��u�ǂ̓ˏo�x���̈Ⴂ���s���ĂɂȂ�B�����|�ɒB���Ă��Ȃ������u�ǂł́A���̓��������[�u���悵�A�אڂ����u�ǂ����킸���ɐ���オ��B���[�u�̓�������́A�����̊u�ǂ͒����������܂��Ⴍ�Ȃ�Ȃ��璆���|����1cm�O��L������B�̗̂��ʂł́A��������1cm�قǂ܂ł͐�[����������������̘]���悭���B���A���̕���������オ��B���̓������璆�S�܂ł͂�≚��ł���A�]�����܂蔭�B�����A�����݂̂��g�ł��Ȃ�����ˏ�ɔz��B�]�ԂɌ�����̕ǂ��k���ŕ\�ʂ����炩�A����1�`5mm�E�[��1mm�قǂ̍ג����E���s�K���Ɍ�����B
�������F��ʌ�g������̎ア�ʎΖʁE��ʎΖʂ̊�Տ�⍻�I��Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�FGittenberger et al. (2011) �ɂ���āA�{��݂̂��܂ޑ��ƂȂ���Lobactis ��Fungia fungites �V�^�U���N�T�r���C�V�i�{�K�C�h���f�ڎ�j�݂̂��܂ޑ��ƂȂ���Fungia �̘a���́A���ꂼ��̃^�C�v��̘a�����Ƃ��āA�N�T�r���C�V���ƃV�^�U���N�T�r���C�V���ɉ��̂���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Podabacia crustacea (Pallas, 1766)
���G���}�J�����T���S |
Fungiidae Dana, 1846 �N�T�r���C�V��
Podabacia Milne Edwards and Haime, 1849 ���G���}�J�����T���S�� |
|

�B�e�F������^(���V�\�s��Óc�A���[17m)

|

䰂̒��a��3�`5mm�قǁB
|
|
�����`�F�t��`��Q�́B�t��E���ߏ�����ɐL�тĂ��蔫��ɂȂ�����A�����Ɋ������܂�ĉ~����ɂȂ�����A��d�ɂ��d�Ȃ����肷�邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�ŁA�Ԃ݂�тт����̂�ΐF�ɋ߂����̂�������B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA䰂̒��a��3�`5mm�قǁB�̂͌Q�̏�ʂ݂̂ɖ��ՂȂ����z���A�̊Ԋu���قړ������B�c�Q�̂ł����S�̂͊m�F�ł��Ȃ��B�̔z��̓T���i�X�e���C�h�^�ŁA�����I�ɓ��S�~��ɔz��B�u�ǁE�]��3���܂Ŕ��B�B1���u�ǁE�]�͌����A�悭�ˏo����B1���u�ǁE�]�̏㉏�ɂ͑傫���̑��������[�u��̋������A���̋����̏㉏�ɂ͏����Ȟ���̑��������ꂼ��悭���B����B����ČQ�̕\�ʂ̓U���U�����Č�����B�������A�悭�ΊD�������Q�̂ł́A���[�u��̋��������܂�ڗ����Ȃ��B2����3���̊u�ǁE�]�́A1���u�ǂɔ�ׂĔ����ˏo�����A���[�u��̋����⋘����̑����̔��B�͈����B�����͖_��A�܂��͕����̖_��̂��̂����ɕ���ŗZ��������ŁA�悭�ΊD�������Q�̂ł͖��Ă����A�ΊD�����ア�Q�̂ł͕s���āB1���E2���u�ǂ͎����܂ŒB���邪�A3���u�ǂ͒B���Ȃ��B�����͐��������ɐL�т���̘]�Ɛ��������ɐL�т��_��̃V�i�v�e�B�L��������Ȃ�B�]�Ԃ̃V�i�v�e�B�L�����́A�]�̉����œ��Ԋu�ɋK���������z��B䰕ǂ��܂����l�̃V�i�v�e�B�L�����Ō`�������B�Q�̗��ʂɂ���ʂɑΉ�����]�����B���邪�A��ʂɔ�ׂĕs���āB�]�̏㉏�ɂ́A����̑��������������`������邪�A�傫����Ԋu�͕s�����B�]�Ԃ̋����ɂ͕s�K���J�����E�����S�~��ɕ��Ԃ��Ƃ�����B
�������F�g������̎ア�ʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Polyphyllia talpina (Lamarck, 1801)
�C�V�i�}�R |
Fungiidae Dana, 1846 �N�T�r���C�V��
Polyphyllia Quay and Gaimard, 1833 �C�V�i�}�R�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[20m)
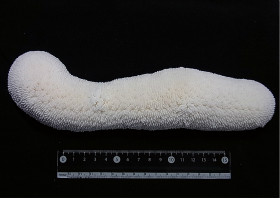
�������ʐ^�̌Q�̂Ƃ͕ʌQ��(���ꌧ�Ί_���Ɍ��Ԙp�A���[20m�ō̏W) |
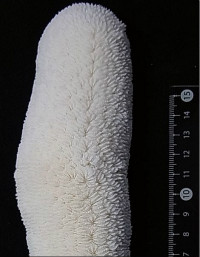
|
|
�����`�F�Q�͔̂�Œ����B�O�`�͎�ɝG���ȑȉ~�`�ŁA�����ɔ����ČQ�̒��������ʌ^�ɐ���オ���Ă���B���a�����\cm�ɂȂ�܂Ő�������B
��̕��̐F�ʂƓ����F���F�B���Ԃł��A�Q�̕\�ʂ��疞�ՂȂ��G���L���Ă���B�G��̐�[�ׂ͍��A�킸���ɔ����B
���i�̓����F�̂͑��p�`�ŁA���a1cm�O��̂��̂��قƂ�ǁB�̔z��͊�{�I�ɃT���i�X�e���C�h�^�B�ȉ~�`�Q�̂ł́A����オ�������S���̒��������ɕ����̌̂�������ɕ��сA����ɂ��̍��E�ɕ����̌̂��قڕ��s�ɕ��邱�Ƃ�����B�u�ǂ�3���܂Ŕ��B����B���ɌQ�̎����̌̂ł́A�������̊u�ǂ��悭�L�����A�قڕ��s�ɔz��B�u�ǂ̏㉏�Ƒ��ʂɂ́A������̑������������ƓˋN�����ꂼ�ꔭ�B����B���̂��߁A�אڂ���u�NJԂ͋���������B1���E2���u�ǂ͎����ɒB����B1���u�ǂ�䰕Ǐ㉏�Ŕ��Ɍ����A����ɂ悭�ˏo���邪�A䰓��ł͒����������A�ˏo���邱�Ƃ��Ȃ��B�܂��̂ɂ���ẮA1���u�ǂ��Z���ԕُ�ɂȂ邱�Ƃ�����B2���u�ǂ͔����A䰕Ǐ㉏���玲���߂��܂łقړ��������B3���u�ǂ͔��B�������A�����ɒB���邱�Ƃ͂Ȃ��B������1���E2���u�Ǔ����̃p����t�Ɨאڂ���u�NJԂ��Ȃ��g���x�L�����Ō`������邪�A���S�ɖ��ĂȖ_��ˋN�Ȃǂ͌����Ȃ��B�Q�̗��ʂɂ͘]���`������邪�A�������邢�͘]���̂����ʂ��邱�Ƃ�����Ȃ��Ƃ������B�]�̏㉏�ɂ́A�傫�����悭�����A��[��������̑��������������悭���B����B�̕ǂ��k���ŁA�s�K���ɍE��������B
�������F�g������̎ア�ʎΖʂ��ʎΖʂ̍��I��Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Acanthastrea echinata (Dana, 1846)
�q���I�I�g�Q�L�N���C�V |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Acanthastrea Milne Edwards and Haime, 1848 �I�I�g�Q�L�N���C�V�� |
|

�B�e�F�����C��(����q���n���̊≮�A���[7m)
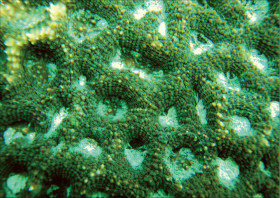
|

䰂̒��a��8mm�`2cm�B
|
|
�����`�F�핢��`�����Q�̂ŁA��Ղ��L�������Ȃ��璷�a20cm�ȏ�̑傫�ȌQ�̂ɐ�������B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�B���Ղ����`�D�F�����������̂������B��͓̂��������A�u�Ǐ㉏�ɐ��R�ƕ��������ˏo����l�q���悭������B�����ɂ���ĉ����グ��ꂽ��̊O�炪�A�����̓��S�~���ᰂƂȂ��Č����邱�Ƃ�����B䰕ǂ͌����A䰂͐[���A�̌��Ղ̌`��傫���͑����Č�����B�����A�̊Ԃ̋��E�͕s���āB
���i�̓����F�͉̂~�`�܂��͊p�����~�`�`���p�`�ŁA䰂͒��a8mm�`2cm�ŁA1cm�O��̂��̂������B�̔z��̓v���R�C�h�^�`�Z���I�C�h�^�ŁA�o��r���Ō̂�2�`4�Ȃ������ӏ����H�Ɍ�����B䰕ǂ͌���2�`4mm�B�u�ǁE�]��4���܂Ŕ��B�A�����ɂ�炸���������A���Ԋu�ł��ꂢ�ȕ��ˏ�ɔz��B������䰕Ǐ㉏�ł��܂�ˏo�����A䰐S�Ɍ������ċ}�X�ŗ������ނ��߁A䰂͐[��������B䰂̒��a��1cm�قǂ̌̂Ŋu�ǂ�30�����x�B�אڌ̂ŁA�����]�����L����u�ǂ̎����͈�v���Ȃ��Ƃ��낪�����B1���E2���u�ǂ͎����܂ŒB����B�ʏ�3���u�ǂ͎����܂ŒB���Ȃ����A�������悭���B�����̂ł͒B���Ă���悤�Ɍ�����B4���u�ǂ͒Z���A1���E2���u�ǂ̔������x�̒����B�u�ǁE�]�̏㉏�ɂ́A��[�����[�u��܂��͐�����������قړ��Ԋu�Ŕz��B���ɁA䰕Ǐ㉏�ɋ߂�2�`4�̋����͊���悭������A��[������ď���ɂ悭�ˏo����B���̂��߁A�u�ǁE�]��䰕Ǐ㉏�̂ق�������������B�u�ǁE�]�Ƌ����̑��ʂɂ́A������������̑������悭���B����B�����͒��a2�`4mm�̉~�`�`�ȉ~�`�Ŗ��ĂȂ��̂��قƂ�ǁB
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Acanthastrea hemprichii �q���^�I�I�g�Q�L�N���C�V�ƍ�������Ă���A�����ł̐������Č�������K�v������B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FA. lordhowensis�Ƃ��ċL�^�����\������
|
|
| |
Acanthastrea hemprichii (Ehrenberg, 1834)
�q���^�I�I�g�Q�L�N���C�V |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Acanthastrea Milne Edwards and Haime, 1848 �I�I�g�Q�L�N���C�V�� |
|

�B�e�F�����C��(����q���n���̊≮�A���[12m)
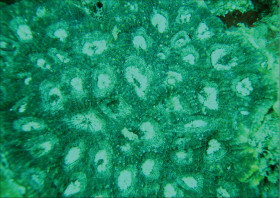
|

䰂̒��a��8mm�`1.4cm�B
|
|
�����`�F��ɔ핢��Q�́B�Q�̂͒��a20cm���邱�Ƃ͏��Ȃ��B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�B���Ղ����`�D�F�̂��̂������B�Ԃ≩�ȂǑN�₩�Ȃ��̂�������B��̂͂����������A䰕Ǐ㉏�ł͊u�Ǐ㉏�̋��������ʂł���B�����ɂ���ĉ����グ��ꂽ��̊O�炪�A�����I�ɓ��S�~���ᰂƂȂ��Č����邱�Ƃ�����B䰕ǂ͌����A䰂͐�����B�����A�̊Ԃ̋��E�͕s���āB
���i�̓����F�̂͊p�����~�`�`���p�`�ŁA䰂͒��a8mm�`1.4cm�A�ʏ��1cm�O��̂��̂������B�̔z��͂قڃZ���I�C�h�^�A�o��r���̌̂������Ȃ��邱�Ƃ͋H�B䰕ǂ͔����A1�`2mm�B�u�ǂ�4���܂Ŕ��B���A�����ɂ�炸�قړ����������A1���u�ǂ��������B�u�ǂ�䰕Ǐ㉏�ł��܂�ˏo�����A䰐S�Ɍ������Ċɂ₩�ɌX���邽�߁A䰂͐�����B䰂̒��a��1cm�قǂ̌̂Ŋu�ǂ�30�����x�B�אڌ̂ŁA�����]�����L����u�ǂ͎�������v����Ƃ��낪�����B1���E2���u�ǂ͎����܂ŒB����B3���u�ǂ͒ʏ펲���ɂ͒B���Ȃ����A�����̂�����O�܂ŐL�т�B4���u�ǂ�1���E2���u�ǂ̔������x�̒����B�u�ǁE�]�̏㉏�ɂ́A�~����Ő�[��������������قړ��Ԋu�Ŕz��B���ɁA䰕Ǐ㉏�ɋ߂�2�`4�̋����͏���������Ă���A�s�K���ɔ�����邱�Ƃ�����B�u�ǂƋ����̑��ʂɂ�������̑����������邪�A�܂�Ŗڗ����Ȃ��B�����͒��a3mm�����̉~�`�`�ȉ~�`�ɂȂ邪�A�s���Ă܂��͌������Ƃ������B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Acanthastrea echinata �q���I�I�g�Q�L�N���C�V�ƍ�������Ă���A�����ł̐������Č�������K�v������B
** ����̒����ł͊m�F�ł��Ȃ������������EVeron(1995)�ł͎�q���Ŋm�F����Ă����
|
|
| |
Acanthastrea hillae Wells, 1955
�I�I�g�Q�L�N���C�V |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Acanthastrea Milne Edwards and Haime, 1848 �I�I�g�Q�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��Óc�A���[10m)

|

䰂̒��a��2.5�`4.5cm�B
|
|
�����`�F�핢��`�����Q�̂ŁA���a10�`20cm�قǂ̏����߂̌Q�̂������B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�ŁA�D�F�����������̂������B�Q�̕\�ʂ��s�K���Ȃ܂���͗l�ɂȂ�����A���Ղ܂��͊u�Ǐ㕔�����ƈقȂ�F�ʂɂȂ����肷�邱�Ƃ�����B��͓̂����ŁA���̕\�ʂ������Ɍ�������̂�A䰕Ǖt�߂�����Ɍ�������̂�����B䰕Ǐ㉏�ł́A�̊Ԃ̋��E�����Ăȕ����ƕs���Ăȕ��������݂���B
���i�̓����F�͍̂ג������p�`�`���p�����~�`�ŁA䰂͒��a2.5�`4.5cm�A3cm�قǂ̑��p�`�������B�̔z��͎�ɃZ���I�C�h�^�����A�����ȌQ�̂ł̓v���R�C�h�^�ɂȂ�A�傫�Ȓ��S�̂����ʂł���B�傫�ȌQ�̂ł́A���S�~��Ɍ̂��A�Ȃ��āA����10cm����J���`������邱�Ƃ�����B䰕ǂ͒����������قړ������������A����Q�̓��ł������͕ς��₷���A�Z���I�C�h�^�Q�̂�1.5�`5mm�قǁA�v���R�C�h�Q�̂ł�1cm�߂��ɂȂ邱�Ƃ�����B�u�ǂ͒ʏ�4���܂Ŕ��B���A�傫�Ȍ̂ł͒Z��5���u�ǂ�䰕Ǐ㉏�t�߂Ɍ`������邱�Ƃ�����B�u�ǂ̌����͂قړ��������A1���u�ǂ����̊u�ǂ��������Ɍ����Ȃ邱�Ƃ������B�v���R�C�h�^�Q�̂ł́A䰕NJO���̊u�lj������ŒZ���]���`������邪�A�u�ǂƂ̋�ʂ͓���B1�`4���u�ǂ́A䰕Ǐ㉏��������Ɍ������}�X�ŗ������ނ��A�r������X�͊ɂ₩�ɂȂ�B1�`3���u�ǂ͎����ɒB����B4���u�ǂ͐L�тĂ�3���u�ǂ�3/4�قǂŁA���̓����͘p�Ȃ��Ȃ���3���u�ǂɗZ������B�u�ǁE�]�̏㉏�ɂ́A��[�����A�傫�����悭�������������悭���B���A���R�ƕ��ԁB�܂�䰕Ǐ㉏�`���������̋����͑傫���A����Ɍ������Ă悭�ˏo����B�u�ǁE�]�Ƌ����̑��ʂɂ́A������������̑������悭���B����B�����͒��a2.5�`5.5mm�̉~�`�`�ȉ~�`�Ŗ��āB
�������F���Օ��I�Ȋ�ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʓ��ȓ�B��q���ł͋H�B
�⑫�F�{���Acanthastrea ishigakiensis �C�V�K�L�I�I�g�Q�L�N���C�V��A. bowerbanki �i����Ƃ��ɖ{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Acanthastrea cf. lordhowensis Veron and Pichon, 1982
�J�N�I�I�g�Q�L�N���C�V |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Acanthastrea Milne Edwards and Haime, 1848 �I�I�g�Q�L�N���C�V�� |
|

�B�e�F�[���T�L(���V�\�s�Y�c�p�A���[5m)

|

䰂̒��a��8mm�`1.3cm�B
|
|
�����`�F�핢��`�����Q�́B���a5�`15cm�قǂ̏����Ȃ��̂������B
��̕��̐F�ʂƓ����F��ɒW�ΐF��W���F�ŁA�ԐF�Ȃǂ̑N�₩�ȐF�ʂɂȂ邱�Ƃ�����B�Q�̕\�ʂ��s�K���Ȃ܂���͗l�ɂȂ�����A䰓��`�̋��E�����S�~��ɈقȂ�F�ʂɂȂ�����A䰕Ǐ㉏�Œ�u�Ǒ������������Ȃ����肷�邱�Ƃ�����B��͓̂��������A�Q�̕\�ʂ̓U�����Č�����B�����A�̊Ԃ̋��E�͖��āB
���i�̓����F�̂͑��p�`�ŁA䰂͒��a8mm�`1.3cm�A���S�ȃZ���I�C�h�^�z��B䰕ǂ͌���1�`2mm�ŁA��̂ق��������߁B�u�ǂ�4���܂Ŕ��B���邪�A4���u�ǂ̔��B�������̂������B䰂̒��a��1cm�قǂ̌̂Ŋu�ǂ�26�`36�����x�B��{�I��1�`3���u�ǂ͓��������ŁA䰕Ǐ㉏�œˏo���A䰐S�Ɍ������Ċɂ₩�ɌX����B������1���E2���u�ǂ́A䰕Ǐ㉏�ŕs�K���ɔ�����Ă���ɓˏo���邱�Ƃ�����B4���u�ǂ͒Z���A�����L�тĂ����̊u�ǂ̔������x�܂ŁB�u�Ǐ㉏�̋����͑傫����Ԋu���s���������A䰕Nj߂���2�`3�͏���Ɍ������Ă悭�ˏo����B�����̈ꕔ���~����ɔ�����āA�s�K���ɓˏo���邱�Ƃ�����B�u�ǂƋ����̑��ʂɂ�������̑����������邪�A�܂�Ŗڗ����Ȃ����Ƃ������B�����͉~�`�`�ȉ~�`�ŁA���a2�`3.5mm�Ƒ傫�߁B����Q�̓��ł����ĂȂ��̂���s���ĂȌ̂܂ŗl�X�B
�������F���Օ��I�Ȋ�ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Acanthastrea lordhowensis �i�{�K�C�h���f�ڎ�j�Ɠ��肳��Ă����B����������ɂ́A�̂̑傫���A�u�ǂ̔��B�E�z��l���A�����̌`��ȂǑ����̌`�ԓI�����̈Ⴂ���F�߂���B����āA����X�Ȃ镪�ފw�I�������K�v�Ǝv����B�܂��{��́AA. aff.lordhowensis �Ƃ���������Ă��邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Acanthastrea aff. lordhowensis Veron and Pichon, 1982
�}���I�I�g�Q�L�N���C�V�i�V�́j |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Acanthastrea Milne Edwards and Haime, 1848 �I�I�g�Q�L�N���C�V�� |
|

�B�e�F�����@�O(���茧���s�Y�A���[2m)

|

䰂̒��a��1.5cm�قǁB
|
|
�����`�F�����`���Q�́B���a20cm����h�[���^�`�����`�̌Q�̂��`������B
��̕��̐F�ʂƓ����F��ɒW�ΐF��W���F�B䰕Ǐ㉏�̒�u�ǁE�]�̑������������Ȃ�A�Q�̕\�ʂ��s�K���Ȃ܂���͗l�Ɍ����₷���B��͔̂��ɓ����ŁA�G��Ă��Ȃ��Ȃ��k�܂Ȃ��B�̋��E�ɂ́A���D���t�����邱�Ƃ�����B�����A�̊Ԃ̋��E�͖��āB
���i�̓����F�͉̂~�`�`�p�����~�`�B�̂����W����Ƃ���ł͂��тȌ`�ɂȂ�₷���B䰂͒��a1.5cm�قǂŁA�o��r���̂��͍̂ג���3cm�߂��ɂȂ邱�Ƃ�����B�̔z��́A�Q�̏���ł̓Z���I�C�h�^�����A�����ł̓v���R�C�h�^�ɂȂ�₷���B�אڌ̂̋��E�ɂ́A���їނ̊Ǐ�̐��ǂ����B���邱�Ƃ�����B䰕ǂ͌Q�̏����2�`3mm�A�����ł�4mm�߂��ɂȂ�B�u�ǁE�]�͒ʏ�4���܂Ŕ��B���A�傫�Ȍ̂ł�䰕Ǐ㉏�ɒZ��5���u�ǂ��`������邱�Ƃ�����B�u�ǂ�䰕Ǐ㉏�œˏo���Ȃ����悭������A�X�͊ɂ₩�B䰌a1.5cm�قǂ̌̂Ŋu�ǂ�40���O��B1���E2���u�ǂ͑��̊u�ǂ����������A�����܂ŒB����B3���u�ǂ͎�����O�܂ŐL�т邪�����ɂ͓͂����A�p�Ȃ��Ȃ���2���u�ǂɗZ������B4���u�ǂ�1���E2���u�ǂ̔������x�̒����B�]�͒Z�����A�u�NJO���Ɠ��l�ɂ悭�������B�u�ǁE�]�̏㉏�ɂ́A�傫�����������ݒ[�ő����~����̋������A����������Ȃ���L���Ԋu�Ŕz��B���ɂ́A��[��������Ğ��_��ɂȂ���̂�����B�u�ǁE�]�Ƌ����̑��ʂɂ́A������������̑������悭���B����B������2�`5mm�Ƒ傫���B
�������F�g�Q�̉e���̏��Ȃ���ʎΖʂ̐��[5m�Ȑ[��A�Օ��I�Șp�̐��[5m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧�Δn�ȓ�B��q���ł͊m�F����Ă��Ȃ��B
�⑫�F�{���Acanthastrea cf.lordhowensis �ƍ�������Ă���A�����ł̐����̍Č������K�v�B�V�̘a���́A�v���R�C�h�^�ɔz��~�`�̂������邱�ƂɗR������B�{��̐����ʐ^�ƍ��i�ʐ^�̌Q�͈̂قȂ邪�A�B�e�n�_�ƍ̏W�n�_�͂قړ����B
* ��q�����L�^��

|
|
| |
Cynarina lacrymalis (Milne Edwards and Haime, 1848)
�R�n�i�K�^�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Cynarina Brüggemann, 1877 �R�n�i�K�^�T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[13m)

|

�u�ǂ̋����̓��[�u��B
|
|
�����`�F�Œ����̒P�̂ŁA�~����`�ȉ~����B�����H�ɁA5�`6�̂��t�@�Z���C�h�^�ɔz���ԑ���Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɒW���F�`�W�ΐF�B�N�₩�ȃs���N�`�Ԃ݂����������̂�����B�|���v�͒��Ԃł��L�тĂ���A�������̊O�������X�̊u�ǂ��ݍ��ނ悤�ɖA��ɂȂ�����A���D�̂悤�Ɍ̑S�̂��ݍ��ނ悤�ɖc��肷��B��҂ł́A���Ղ𒆐S�ɕ����̔������䂪���S�~��Ɍ����邱�Ƃ�����B
���i�̓����F�̂͑ȉ~�`���Z�����������тȑȉ~�`�B�O�҂ł�䰂̒��a�͍ő��7cm�قǁA��҂ł�9cm�قǂɂȂ�B�u�ǁE�]��5���܂Ŕ��B���A䰕Ǐ㉏�Ō����悭�ˏo����B�u�ǂ̌�����䰕Ǐ㉏�ł̓ˏo�x���͎������Ƃɖ��ĂɈقȂ邪�A�]�͌����������������ɂ�炸�قړ����B�u�ǂ̌X�͊ɂ₩�ŁA1�`3���u�ǂ͎����܂ŒB����B4���E5���u�ǂ͎����ɂ͒B�����A4���u�ǂ͎����̂���O�ŁA5���u�ǂ͂���ɂ��̎�O�ŁA�p�Ȃ��Ȃ���3���E4���u�ǂɗZ������B�u�Ǐ㉏�ɂ̓��[�u��̑傫�ȋ������`������A��u�ǂقǑ傫���Č����B�����̌`���Ԋu�́A1�`3���u�ǂł͕s���������A4���E5���u�ǂł͂悭�����B����A�]�̋�����䰕Ǐ㉏�߂��ł悭���B����B�����͎����ɂ�炸�قړ����傫���ŁA��[�������A�傫�����Ԋu���悭�����B�u�ǁE�]�̕\�ʂɂ́A������̑������т�����Ɣ��B����B�����͑ȉ~�`�ő傫���A䰂̒��������̒����̖�1/3���߂�B�����́A�u�Ǔ�������L�т������̎���̃g���x�L���������W���Č`������̂��悭������B
�������F���D���͐ς���悤�ȏʎΖʂ��ʎΖʂ̐��[20m�Ȑ[�Ō�����B�J���I�Șp�ł͂�������Ƃ���ŁA�Օ��I�Șp�ł͐��[10m�Ȑ�ł�������B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Echinophyllia aspera (Ellis and Solander, 1786)
�L�b�J�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Echinophyllia Klunzinger, 1879 �L�b�J�T���S�� |
|
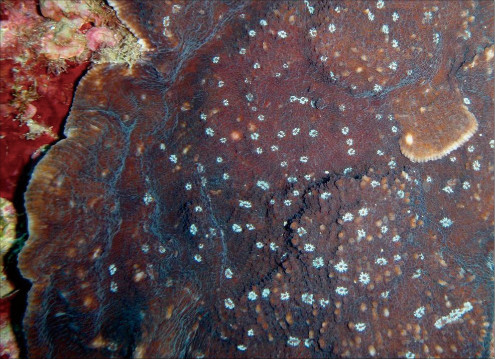
�B�e�F�쑺�b��(����q���n���̊≮�A���[8m)

|

䰕ǂƎ����̔��B�͈����B
|
|
�����`�F�핢��`�t��Q�́B�����H�ɗt�������オ��A��d�ɂ��d�Ȃ����Q�̂������邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�ΐF�B�Q�̑S�̂��N�₩�ȐԐF�≩�ΐF�ɂȂ邱�Ƃ�A���Ղ̐F�ʂ����ƈقȂ邱�Ƃ�����B�̂͂ǂ�����݂��Ă���A�����I�ɓ��S�~��z��Ɍ����邱�Ƃ�����B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA�Q�̎����̌̂�䰂̒��a��5mm�`1cm�قǁA�o��r���̌̂͂���ȏ�̑傫���Ɍ�����B�����ȌQ�̂ł́A���a1.5cm�قǂ̒��S�̂������邱�Ƃ�����B�����̂�䰕ǂ������A���������ɖ��݂܂��͂킸���ɐ^��ɓˏo����B�����̂̊u�ǁE�]�͒ʏ�3���܂Ŕ��B����B1���E2���u�ǁE�]�́A3���u�ǁE�]�ɔ�ׂ�ƌ����A�悭�ˏo����B�܂��A1���u�ǂ̂ق���2���u�ǂ����������B1���E2���u�ǂ͎����܂ŒB���邪�A3���u�ǂ͎����܂ŒB���Ȃ��B2���E3���̊u�ǂƘ]�̓����ɂ́A䰂̕������܂߂ċ��������ɉ��݂��`������邱�Ƃ����邪�A�����ɍE���J�����Ƃ͂Ȃ��B�u�ǁE�]�̏㉏�ɂ́A�ו���[�����������قړ��Ԋu�Ŕ��B����B�������Q�̗��ʂł́A�T���ċ����̔��B�������B�����͒��a2�`4mm�قǂ̑ȉ~�`�ɂȂ邪�A�����̔��B�������̂��悭������B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B���D���͐ς��₷���Օ��I�Șp�ł́A���[5m�Ȑ�ł����ʂɌ����邱�Ƃ�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Echinophyllia cf.orpheensis �A�o���L�b�J�T���S�AE. echinata ��E. patula �i����Ƃ��ɖ{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����͍Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Echinophyllia cf. orpheensis Veron and Pichon, 1980
�A�o���L�b�J�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Echinophyllia Klunzinger, 1879 �L�b�J�T���S�� |
|

�B�e�F�����C��(���V�\�s��v�ۍ`�A���[6m)

|

䰕ǂƎ������悭���B����B
|
|
�����`�F��ɔ핢��Q�̂����A�Ζʏ�ł͌Q�̎������t��ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�ΐF�B���Ղ̐F�ʂ����ƈقȂ邱�Ƃ�A�Q�̑S�̂��N�₩�ȐԂ݂≩�F�݂�тт邱�Ƃ�����B�Q�̒��S�t�߂̌̂͌Q�̕\�ʂ���s�K���ɓˏo���A�̊Ԋu���s�����ɂȂ�₷���B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA�Q�̎����̌̂́A䰂̒��a��6�`8mm�قǂɂȂ�B�����ȌQ�̂ł́A���a1.5cm�����̒��S�̂������邱�Ƃ�����B�Q�̎����ł́A䰕ǂ����������ɖ��݂���̂ƁA�̂̔����܂��͑S�̂�䰕ǂ��`������đ����܂��͏���ɓˏo����̂�����B�u�ǁE�]�͒ʏ�3���܂Ŕ��B����B1���E2���u�ǁE�]�́A3���u�ǁE�]�ɔ�ׂ�ƌ����A�悭�ˏo����B�܂��A1���u�ǂ̂ق���2���u�ǂ����������悭�ˏo����B1���E2���u�ǂ͎����܂ŒB���邪�A3���u�ǂ͂�����1/2�`2/3�̒����ŁA�����܂ŒB���Ȃ��B2���E3���̊u�ǂƘ]�̓����ɂ́A䰂̕������܂߂ċ��������ɖ��Ăȉ��݂��`������邪�A�����ɍE���J�����Ƃ͂Ȃ��B�u�ǁE�]�̏㉏�ɂ́A�ו���[�����������悭���B���A����䰕Ǐ㉏�t�߂�1���E2���u�ǁE�]�ł͂悭������ēˏo����B�������Q�̗��ʂł́A�����͂قƂ�nj`������Ȃ��B�����͖��ĂŁA���a2.5�`5mm�قǂ̑ȉ~�`�ɂȂ�B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō����邪�A�J���I�Șp�ł͐��[5m�Ȑ�ł�������B�܂��A�ʒr���ʌ̊�Տ�ł��H�Ɍ����邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���ȓ�B��q���ł͋H�B
�⑫�FEchinophyllia orpheensis �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�́A�̂�1�`2cm�قǕs�K���ɍ����ˏo����B�܂�䰂̒��a�͂ǂ��1cm���A�u�ǁE�]�͑S�̓I�ɂ悭�������B����āA����܂ō����ŃA�o���L�b�J�T���S�Ƃ��Ă����{��Ƃ͕ʎ�̉\���������B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Lobophyllia corymbosa (Forskål, 1775)
�}���n�i�K�^�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Lobophyllia de Blainville, 1830 �n�i�K�^�T���S�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[10m)

|

䰕ǂ̓��b�p��ɏ��������
|
|
�����`�F�h�[���^�`�����`�̉ԑ���Q�́B���a1m���邱�Ƃ��������Ȃ��B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�����A�Â��F�ʂ̂��̂��疾�邢�F�ʂ̂��́A���Ղ̂ݐF�ʂ��قȂ���̂ȂǗl�X�B�̂̐�[�́A��₷�ڂ悤�Ɍ�����B
���i�̓����F�̂͂��тȑȉ~�`����ג����ȉ~�`�ŁA䰂͒��a2�`4cm�ŁA3.5mm�O��̂��̂������B�������A�o��r���Ŏ������Q�`3�A�Ȃ����̂ł́A�Z������䰕ǂ��s�K���ɉ����L���^��ԕُ�ɂȂ�A���a��4�`6cm�قǂɂȂ邱�Ƃ�����B�̔z��͊�{�I�Ƀt�@�Z���C�h�^�ŁA������3�ȏ�A�Ȃ��Ă��t���x�����A���h���C�h�^�ɔz�邱�Ƃ͂Ȃ��B�אڂ���}�̊Ԋu�͂قړ����ŁA�݂���7mm�`1cm���x�����B䰕ǂ�2�`3mm�ŁA����Ɍ������ă��b�p��ɊJ���Ă���A�����I�ɑ����ɂ߂���邱�Ƃ͏��Ȃ��B�u�ǁE�]��5���܂Ō`������A��̂��̂قnj����A�ˏo����B�u�ǁE�]�̏㉏�ɂ͋������悭���B���A䰕Ǐ㉏�ʼns������Ă悭�ˏo����B�܂��A�u�Ǐ㉏�̋����͏�����A�]�㉏�̋����͏�����瑤���������₷���B�����̋����̑傫���́A�]�̂��̂����u�ǂ̂��̂��傫��������B�����́A���a4�`8mm�قǂ̉~�`�`�ȉ~�`�Ŗ��āB
�������F�ʒr���ʌ���ʎΖʂ̗l�X�Ȋ��ɐ������邪�A��������g�Q�̉e���̏��Ȃ����ő���������B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Lobophyllia hemprichii �I�I�n�i�K�^�T���S�ƍ�������Ă���\�������邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F���iAcanthastrea amakusensis�Ƃ��ċL�^�j
|
|
| |
Lobophyllia hemprichii (Ehrenberg, 1834)
�I�I�n�i�K�^�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Lobophyllia de Blainville, 1830 �n�i�K�^�T���S�� |
|

�B�e�F�����C��(���V�\�s�Y�c�p�A���[8m)

|

�]�㉏�ł��������悭���B����B
|
|
�����`�F�h�[���^�`�����`�̉ԑ���Q�̂ɂȂ�B�Q�̂̒��a��1m���邱�Ƃ��������Ȃ��B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�B�Ԗ���тт����F��D�ΐF�̂��̂�������B�u�Ǐ㉏�̋����̑���������̈Ⴂ�ɂ���āA�O�ς��傫���قȂ��Č�����B
���i�̓����F�̂͂��тȑȉ~�`�`�ג����ȉ~�`�A���a2�`4cm�A�̂�2�`3�A�Ȃ����t�@�Z���C�h�^��A4�ȏオ�s�K���Ɏ֍s���Ȃ���A�Ȃ��Ē���10cm�ȏ�ɂȂ����t���x�����A���h���C�h�^�̌̔z������B�אڂ���}�̊Ԋu�͂قړ����ŁA�݂���7mm�`1cm���x�����B䰕ǂ�4�`8mm�قǁA�V���Ȍ̂��o�肷��Ƃ���ł́A��⑤���ɂ߂��ꂽ�悤�Ɍ�����B�u�ǁE�]��5���܂Ō`������A��̂��̂قnj����A�ˏo����B�������ΊD�����ア�Q�̂ł́A�����̈Ⴂ��1�`3���u�NJԂ�4���E5���u�NJԂŕs���ĂɂȂ�₷���B�u�ǁE�]�̏㉏�ɂ͋������悭���B���A䰕Ǐ㉏�ł͉s������Ă悭�ˏo����B�܂��A�u�Ǐ㉏�̋����͏�����A�]�㉏�̋����͏�����瑤���������₷���B�����́A���a5�`9mm�قǂ̉~�`�`�ȉ~�`�Ŗ��āB
�������F�ʒr���ʌ���ʎΖʂ̗l�X�Ȋ��ɐ������邪�A��������g�Q�̉e���̏��Ȃ����ő���������B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{���Lobophyllia robusta �n�i�K�^�T���S��L. corymbosa �}���n�i�K�^�T���S�ƍ�������Ă��邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Lobophyllia robusta Yabe, Sugiyama and Eguchi, 1936
�n�i�K�^�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Lobophyllia de Blainville, 1830 �n�i�K�^�T���S�� |
|

�B�e�F���{�@��(���V�\�s���Y�A���[10m)

|

�]�㉏�ł͋����̔��B�������B
|
|
�����`�F�Q�̂������ȂƂ��͔핢��`�����ŁASymphyllia�_�C�m�E�T���S���ƊԈႢ�₷���B�Q�̂̐����ƂƂ��ɁA���ꂢ�Ȕ����`�̉ԑ���Q�̂ɂȂ�B
��̕��̐F�ʂƓ����F�F�ʂ͒����F�A�ԐF��ΐF�ȂǗl�X�B�u���F���������̂�Q�̕\�ʂ��܂���͗l�ɂȂ���̂�����B��͓̂��������A��u�ǂ̏�����s�K���ȓ_����ɐ���オ��B
���i�̓����F�̂̒��a4cm�O��A����炪�A�Ȃ��Č̔z��̓t�@�Z���C�h�^����t���x�����A���h���C�h�^�ɂȂ�B�����ȌQ�̂ł̓��A���h���C�h�^�ɂȂ�₷���A䰕ǂɑ�������R�����͌���4�`8mm�A����2�`3cm���x�B����R�����㉏�̊Ԋu��2�`3cm���x�ɂȂ�B�R���������ɂ́A�傫�ȍa���`������邱�Ƃ������B����A���̑��̔z������傫�ȌQ�̂ł́A�ׂ荇�����}�������I�ɗZ��������A���a1cm�O��A����2.5cm�قǂ̃����e�B�N�����`�������肷�邱�Ƃ�����B�u�ǂ�5���܂łŁA�����Ⓑ���A�R�����㉏�ł̓ˏo�x���͎������Ƃɂ悭�����B����1���u�ǂ͑��̊u�ǂɔ�ׂČ����Ɍ����A2mm�ȏ�ɔ������̂ł悭�ڗ��B�u�Ǐ㉏�ɂ͐�[���ۂ݂�тт��傫�ȋ��������B���邪�A�]�ł͔��B�������B�����͉~�`�`�ȉ~�`�A���a4�`8mm�ő傫�����A�Q�̎����ł͕s���Ă܂��͌����̂�����������B
�������F�g�Q�̉e���̏��Ȃ���ʎΖʂ̐��[5�`20m�Ō�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{��̃^�C�v�Y�n�͍��m���y�������s�O��B�{���Lobophyllia hemprichii �I�I�n�i�K�^�T���S��Symphyllia valenciennesi �i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����͍Č������K�v�B�n�i�K�^�T���S�̘a����S���̂�S. valenciennesi �ł͂Ȃ��{��ł���B
* ��q�����L�^��

|
|
| |
Micromussa amakusensis (Veron, 1990)
�A�}�N�T�I�I�g�Q�L�N���C�V |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Micromussa Veron, 2000 �R�I�I�g�Q�L�N���C�V�� |
|

�B�e�F���{�@��(���V�\�s��v�ۍ`�A���[3m)

|

�̂̒��a��8mm�O�オ�����B
|
|
�����`�F�핢��Q�́B�Q�̂͒��a10cm�����̏����Ȃ��̂��������A���I��╽�R�Ȋ�Տ�ł�20cm���邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�B�N�₩�ȐԐF��ΐF�A�D�F�ɂȂ邱�Ƃ�����B䰕Ǐ㉏��אڌ̂̋��E�͔����ۂ��Ȃ邱�Ƃ������B��͓̂����A���Ղ͍L�������A�Q�̕\�ʂ̓U�����Č�����B�אڌ̂̋��E�͖��ĂŁA�̊Ԃ��悭���ꂽ�Q�̂ł́A�אڌ̂̋��E�ɍ��D���͐ς��Ă��邱�Ƃ�����B
���i�̓����F�̂͑��p�`�`�p�����~�`�ŁA䰂͒��a6mm�`1cm�ŁA8mm�O��̂��̂������B�̔z��̓Z���I�C�h�^�����A�����I�Ƀv���R�C�h�^�ɂȂ邱�Ƃ�����B�̊ԂɁA�Ǐ�܂��͍a��̑��їނ̐��ǂ��s�K���ɔ��B����B䰕ǂ�1mm�O��ō�����2�`3mm�قǁA�����͂�┖���Ȃ�B�u�ǂ�3���܂ł悭���B���A�̂ɂ���ẮA��u�NJԂ�䰕Ǔ����ɓ_����`�؏��4���u�ǂ������邱�Ƃ�����B�v���R�C�h�^�̏ꍇ1�`3���]�����B���邪�A���ɒZ���Ċu�ǂƂ̋�ʂ͍���B1�`3���u�ǂ͔����A���Ԋu�ŋK���������z��B�u�ǂ�䰕Ǐ㉏�ł��܂�ˏo���Ȃ����A�������悭���B���Ĕ�����₷�����߁A���ۂ����悭�ˏo���Č�����B1�`3���u�ǂ̌X�͋}�ŁA1���E2���u�ǂ͎����܂ŒB����B3���u�ǂ͎����ɒB���Ȃ������̎�O�܂ł悭�L�сA���p�Ȃ��ėאڂ����u�ǂ̑��ʂɗZ������B�u�Ǐ㉏�ɂ͐�[�̐���������ȋ������A���ʂɂ�������̔����ˋN���悭���B����B䰕Ǐ㉏�߂��ł́A����ł�������1�`3�̑��������������悭���B���A����ɂ悭�ˏo����B�����͒ʏ�2�`3mm�̉~�`�`�ȉ~�`�����A4mm�قǂɂȂ�䰒�̍L�͈͂��߂邱�Ƃ�����B
�������F���D���͐ς��₷����ʎΖʂ̐��[15m�Ȑ�(����5m�Ȑ�)�̊�ՁE���I��Ō�����B
�����ł̕��z�F�É������ɓ��E���茧��ܓ����ʓ��ȓ�B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͌F�{���V�����僖���B�{�킪������Micromussa �̘a���͐���(2013)�Ɋ�Â��B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Oxypora lacera (Verrill, 1864)
�A�i�L�b�J�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Oxypora Saville-Kent, 1871 �A�i�L�b�J�T���S�� |
|

�B�e�F�����C��(���V�\�s���Y�A���[12m)

|

�u�ǁE�]�̓����ɍE���J���B
|
|
�����`�F�핢��Q�̂ŁA�Q�̎����͗t��ɂȂ�₷���B�����H�ɗt�������オ��A��d�ɂ��d�Ȃ����Q�̂������邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�ΐF�B���Ղ̐F�ʂ����ƈقȂ邱�Ƃ�����B�Q�̎����ł͕��s�ɓ��Ԋu�ŕ��]���悭�ڗ��B���ˏo�����̂Ɩ��݂����̂�������B��҂ŁA���ՂƂ���ȊO�̐F�ʂ������Q�̂ł́A�͎̂��ʂ��ɂ����B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA䰕ǂ������B�����ȌQ�̂ɂ́A䰂̒��a��1cm�قǂ̒��S�̂������邱�Ƃ�����B�Q�̎����̌̂͒��a3.5�`5mm�قǂŏ������A�����\�ʂ���킸���ɐ^��ɓˏo����B�̂̊u�ǁE�]�͒ʏ�2���܂Ŕ��B����B2���u�ǁE�]�̓�����2���]�̊O���͔������A���̑��ł͂ǂ����قړ��������ɂȂ�B���̌����̈Ⴂ�́A�Q�̎����̕\�ʂƗ��ʑS�̂Ō����B1���u�ǂ͎����܂ŒB���邪�A2���u�ǂ͒B���Ȃ��B2���u�ǁE�]�̓����ɂ͋��������ɉ��݂��`������A2���u�ǂ̓����̉��݂��̂�䰂��`������B�����̉������ł́A2���u�ǁE�]�̓����̕Б��܂��͗����ɍג����E���s�K���ɊJ�����Ƃ������B�u�ǁE�]�̏㉏�ɂ́A�ו���[�����������������B���A���ɉ��݂��`�����ꂽ�����̕\�ʂƗ��ʂł悭������B�����͉~�`�`�ȉ~�`�ɂȂ邪�A���a��1.5mm�����Ɣ��ɏ������ڗ����Ȃ��B
�������F���Օ��I�ȏʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō����A��ʌΓ��ł��H�Ɍ�����B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�B��q���ł͋H�B
�⑫�F�{���Echinophyllia aspera �L�b�J�T���S�ƍ�������₷�����߁A�����ł̐����̍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Parascolymia aff. vitiensis (Brüggemann, 1877)
�I�j�A�U�~�n�i�K�^�T���S�i�V�́j |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Parascolymia Wells, 1964 �A�U�~�n�i�K�^�T���S�� |
|

�B�e�F����_�V(���V�\�s��Óc�A���[20m)

|

䰂̒��a��4�`8cm�Ƒ傫���B
|
|
�����`�F��ɌŒ����̒P�́B�J�b�v��܂��͔w�̒Ⴂ�~����ɂȂ�B�����H��5�`6�̌̂��A�Ȃ����v���R�C�h�^�Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�ŁA�\�ʂ��܂���͗l�ɂȂ邱�Ƃ�����B�|���v�͂��Ȃ���������A�悭�ˏo����1���u�ǂƂ��̏㉏�̋����̈ʒu���悭������B�����ɂ���ĉ����グ��ꂽ��̊O�炪�A�����̓��S�~���ᰂƂȂ��Č����邱�Ƃ�����B
���i�̓����F�͉̂~�`�`�ȉ~�`�B�o��r���̌̂ł͒Z�����������тȑȉ~�`�ɂȂ邱�Ƃ�����B䰂͒��a4�`8cm�قǁB�u�ǁE�]��5���܂Ŕ��B���A�u�ǂɔ�ׂ�Ƙ]�͂��Ȃ�Z��������B������䰕Ǐ㉏�Ō����A�悭�ˏo����B�u�ǂ́A���������Ɍ������Ă����Ȃ�B�u�ǁE�]�̌�����䰕Ǐ㉏�ł̓ˏo�x���͎������ƂɈقȂ�A��̂��̂قnj����悭�ˏo����B�u�ǂ̌X�͊ɂ₩�Ȃ��߁A䰂͐�����B1�`3���u�ǂƈꕔ��4���u�ǂ͎����܂ŒB���邪�A5���u�ǂ͎����ɂ͒B�����A�L�тĂ���u�ǂ̔������x�̒����ɂ����Ȃ�Ȃ��B�u�ǁE�]�̏㉏�ɂ͐�[����������~����̑傫�ȋ������`�������B�����́A�u�Ǐ㉏�łقڐ^����������A�]�㉏�ł͐^���܂��͂�≺���������B�u�ǁE�]�̕\�ʂɂ́A������������̑��������B���邪����ł͔F���ł��Ȃ��B�����͑ȉ~�`�Œ��a7mm�`1.2cm�قǂŁA�u�Ǔ�������L�т������̎���̃g���x�L���������W���Č`�������̂��悭������B
�������F�ʏ�͏ʎΖʂ��ʎΖʂ̐��[20m�Ȑ[�Ō�����B
�����ł̕��z�F��q���̂ق��A����������E���ł����z���m�F����Ă���B
�⑫�F�{���Parascolymia vitiensis �A�U�~�n�i�K�^�T���S�i�{�K�C�h���f�ڎ�j��ALobophyllia �n�i�K�^�T���S���̒��Ԃ̗c�̂ƍ�������Ă���\��������B����č����ł̐����ɂ��ẮA����X�Ȃ钲�����K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F���iMontastrea curta�Ƃ��ċL�^�j
|
|
| |
Symphyllia agaricia Milne Edwards and Haime, 1849
�q���N�`�_�C�m�E�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Symphyllia Milne Edwards and Haime, 1848 �_�C�m�E�T���S�� |
|
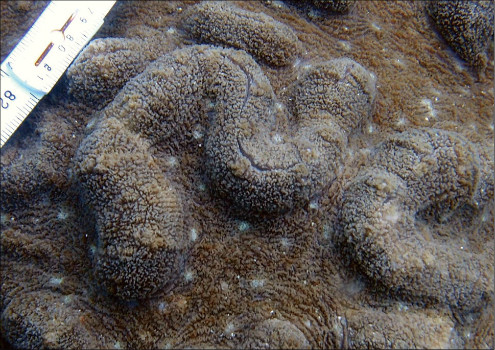
�B�e�F�����C��(���V�\�s�Y�c�p�A���[6m)

|

�R�����㉏�͔���������B
|
|
�����`�F�����`���Q�́B��Ղ��L�������Ȃ��琬�����A50cm���鏀���Q�̂ɂȂ邱�Ƃ�����B���a20cm�ɖ����Ȃ��ꍇ�́A�핢��┼���`�ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B���ΐF��Ԏ��F�Ȃǂ̑N�₩�ȐF�ʂ������̂�����B���Ղ̐F���قȂ邱�Ƃ������B��������ɔ�ׂ�Ɠ����ŁA�Q�̕\�ʂł̋����̓ˏo�͖ڗ����Ȃ��B
���i�̓����F�̔z��͊�{�I�Ƀ��A���h���C�h�^�����A����R�����ԂŎ��������ɕ���A1�̎������̎��������S�~��Ɉ͂ނ悤�ɔz���肷�邱�Ƃ�����B����R�����㉏�̊Ԋu��2.5cm�قǁA������2��ɂȂ��������ł�4.5cm�قǂɂȂ�B�R�����̍�����2�`2.5cm�ƍ����Ȃ�₷���A�����A�Ȃ邱�Ƃ�����A�s�A���ɂȂ��ă����e�B�N�����`������邱�Ƃ�����B�ʏ�A�R�����͌���3�`5mm�����A������1mm�����̔����ɁA���1cm�߂��̌����ɂȂ邱�Ƃ�����A�\�ʂ��s�K���Ɍ�����Q�̂����Ȃ��Ȃ��B�u�ǂ�5���܂Ō`������A��̂��̂قnj����A�悭�ˏo����B�R������ł�4���E5���u�ǂ��`������镔���Ƃ���Ȃ�����������B���̂��߁A������1�`1.5mm�قǂ�1�`3���u�ǂ�0.5mm������4���E5���u�ǂ̔z�K�������������ɂ����B�]�͌Q�̎����`���ʂɂ����Č����邪�A�����̈Ⴂ�͊u�ǂɔ�ׂĕs���āB��u�ǂ̏㉏�ɂ͓ݒ[�ő����̑������������悭���B���邪�A�R���������t�߂ł́A�����̊Ԋu�Ⓑ��������͕s�K���ɂȂ邱�Ƃ�����B�����͉~�`�`�ȉ~�`�ŁA���a4�`8mm�B�傫�ȌQ�̂ł́A�召�̎�����1.5�`2.5cm�̊Ԋu�ŁA�K�����������݂ɕ���ł���悤�Ɍ�����B
�������F�ʎΖʂ��ʎΖʂ̐��[5�`20m�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�{���Symphyllia valenciennesi �i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����̍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Symphyllia radians Milne Edwards and Haime, 1849
�_�C�m�E�T���S |
Lobophylliidae Dai and Horng, 2009 �I�I�g�Q�T���S��
Symphyllia Milne Edwards and Haime, 1848 �_�C�m�E�T���S�� |
|

�B�e�F�i�c�r��(���V�\�s�匴�A���[6m)

|

�R�����㉏�͌���������B
|
|
�����`�F�����`���Q�́B���a40�`50cm���x�̃h�[���^�`�����`�ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�ŁA�Ԃ݂�тт����̂�����B���Ղ̐F�ʂ��قȂ邱�Ƃ������B
���i�̓����F�̔z��̓��A���h���C�h�^�ŁA�����̌̂������A�Ȃ��ăR�������悭�`�������B�R�����͍���1�`1.5mm�ł悭�����A����3�`6mm�Ŋ�̂ق����������B����R�����㉏�̊Ԋu��1.5�`2mm�B�u�ǂ�5���܂Ŕ��B���A䰐S�Ɍ������ċ}�X�ŗ������ށB1�`3���u�ǂ͂قړ���������1mm�O��A�R�����㉏�ł悭�ˏo����B4���E5���u�ǂ͔���0.5mm�O��ŁA�R������ł͓ˏo���Ȃ��B�R�����㉏�ł́A��ƍ����̊u�ǂ����݂ɋK���������z��l�q���悭������B�܂��R���������ł́A�אڌ̂̊u�ǁE�]������悤�ɍׂ��a���s�K���Ɍ`������邱�Ƃ�����B�u�Ǐ㉏�ɂ͋������悭���B���A��̂��̂قǑ傫���A�悭�ˏo����B�����́A�����߂��ł͒Z����[���ۂ����A�R������[�Ɍ������ɂ�Ē����Ȃ�A��[������ď���������₷���B�]�͌Q�̎����̗��ʂɌ����邪�A�����ɂ�������ˏo�x���̈Ⴂ���s���ĂŌ�������̂�����B�܂��A�����̏㉏�ł̋����̔��B�͔��Ɉ����B�����͉~�`�`�ȉ~�`�ŁA���a4�`5mm�A�Q�̎����̌̂������悭���B����B�R�����Ԃŗאڂ��鎲���Ԃ̋�����1.5�`2cm�ł悭�����B
�������F�ʒr���ʌ���ʎΖʂ��ʎΖʂ̗l�X�Ȋ��ɐ�������B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Symphyllia recta �z�\�_�C�m�E�T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����̍Č������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FGoniastrea aspera�Ƃ��ċL�^�����\������

|
|
| |
Astrea curta (Dana, 1846)
�}���L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Astrea Lamarck, 1801 �}���L�N���C�V�� |
|

�B�e�F���{�@��(���V�\�s�Y�c�p�A���[6��)

|

䰌a��4�`6mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̎������Œ���Ղ���V�����邱�Ƃ͋H�B
��̕��̐F�ʂƓ����F���F�`�Ê��F�܂��͗ΐF�B
���i�̓����F�̂͂��ꂢ�ȉ~�`�`���ȉ~�`�ŁA䰌a��4�`6mm�̂��̂��قƂ�ǁB�̔z��̓v���R�C�h�^�ŁA�̂̑傫����Ԋu�͂��s�K���B䰕ǂ͌����A������ɓˏo����B�u�ǁE�]��32���O��ŁA4���܂Ŕ��B����B�u�ǂƘ]�̌����͎����ɂ�炸�قړ����ŁA�㉏�ɐ�[�̐�������������Ԋu�ŕ��ԁB�u�ǂ́A䰕Ǐ㉏�ł������Ȃ邪���܂�ˏo���Ȃ��B�u�ǂ́A䰕ǂ̓����ʂɉ����Ăقڐ����ɗ������݁A䰒��1���E2���u�ǂ݂̂�䰐S�����܂ŐL�тĎ����ɒB����B3���u�ǂ́A䰒��1���E2���u�Ǔ����̂�����O�܂ŐL�т邪�A�����̏ꍇ�����ɂ͒B���Ȃ��B4���u�ǂ́A���̊u�ǂɔ�ׂ�Ɣ��ɒZ���A�������ɐL�т邱�Ƃ͂Ȃ��B�����䰕ǂ̂���������^�ォ�猩��ƁA����1�`3���u�ǂƒZ��4���u�ǂ����݂ɐ��R�ƕ���ł���悤�Ɍ�����B1�`3���u�ǂ̓����ɂ̓p����t�܂��̓p�����`������₷���B�܂����̓����̎����́A�u�Ǔ�������L�т��g���x�L�����ɂ���Č`������Ă���B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�É������ɓ��E���茧�ܓ����]���ȓ�B��q���ł͕��ʎ�B
* ��q�����L�^��

|
|
| |
Caulastraea tumida Matthai, 1928
�^�o�l�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Caulastraea Dana, 1846 �^�o�l�T���S�� |
|

�B�e�F���{ ��(���V�\�s���Y�A���[10m)

|

䰂̒��a��1.5cm�O��B
|
|
�����`�F�ԑ���Q�́B���p�Ȃǂ̎Օ��I�Ȋ��ł́A���a��1m���锼����̑�Q�̂ɂȂ邱�Ƃ��������Ȃ��B�����ȌQ�̂�|���v���悭�c��Q�̂́A����Dipsastraea�L�N���C�V���ƊԈႦ�₷���B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�g���ш�ł͌Q�̑S�̂��D�ΐF�Ȃǂ̖��邢�F�ʂ̂��̂������B�|���v�͓����ŁA��u�Ǐ㉏�͔����Ȃ��ēˏo���Č�����B
���i�̓����F�͉̂~�`�`���тȑȉ~�`�ŁA䰂̒��a��1.5cm�O��̂��̂������B�̔z��͊�{�I�Ƀt�@�Z���C�h�^�B���������Q�̂ł́A�̊Ԃɋ��������B���A�v���R�C�h�^�ɂȂ���̂�����B�u�ǁE�]�͂��ꂼ��30���O��ŁA�ʏ�4���܂Ŕ��B����B���̌����͎����ɂ�炸�قړ����ŁA䰕Ǐ㉏�ł������A�悭�ˏo����B�u�ǁE�]�̏㉏�ɂ́A�u�ǂƒ�������悤�ɂ��G���Ő�[�̐�����������悭���B����B�܂��u�ǂ̑��ʂɂ́A��[�̐���������ˋN���悭���B����B�u�ǂ́A䰕ǂɉ����Ăقڐ����ɗ������݁A���̓����Ƀp����t�͂��܂蔭�B���Ȃ����߁A䰂͐[��������B�����͂悭���B���A�u�Ǔ�������L�т��g���x�L�����Ō`�������B�`�͉~�`�`�ȉ~�`�ŁA���a�܂��͒��a��3�`5mm�قǁB䰕ǂ́A�㕔�ł͔����������ł͂��̓��O�ɖA���g�D���`�������B�Q�̊�̗��ʂ�~����ɐL�т��̂̊O���ʂɖ��ĂȃG�s�e�J�͌`������Ȃ��B
�������F���D���͐ς��₷���Օ��I�ȓ��p��A�ʎΖʂ̐[�݂ȂǂŌ�����B��ʌ̐ł������܂�Ɍ����邱�Ƃ�����B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
* ��q�����L�^��

|
|
| |
Coelastrea sp. 1
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Coelastrea Verrill, 1866 �p���J���m�R�L�N���C�V��(�V��) |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[4m)

|

䰂̒��a��4�`8mm�B
|
|
�����`�F�핢��Q�́B��Ղ��L�������Ȃ��琬������B�Q�̕\�ʂ͕s�K���ɐ���オ��B�Q�̎������Œ���Ղ���V�����邱�Ƃ͋H�B
��̕��̐F�ʂƓ����F���F�`�ΐF�B�u�Ǐ㉏����Ղ������ۂ��Ȃ邱�Ƃ������B䰓��͍L��������B
���i�̓����F�̂͑��p�`�ŁA�����̔z��̓Z���I�C�h�^�B䰂̒��a��4�`8mm�قǂŁA����Q�̏�ł̑傫����`�͂悭�����B�u�ǂ�4���܂Ŕ��B���A䰌a��5mm�قǂ̌̂ł�32���O��B���̏㉏�ɂ́A��[�ɕ����̔����ˋN���������������悭���B����B�����̊u�ǂ͂قړ��������Œ���o�����A䰕Ǔ����ɉ����ċ}�X�ŗ������ށB���̂���䰓��͍L���A�[��������B1���E2���u�ǂ͎����܂ŒB����B�����̊u�Ǔ����ɂ́A6�`10�̃p����t�܂��̓p�����`������A�������͂ނ悤�Ɋ���ɕ��ԁB3���u�ǂ͒ʏ�1���E2���u�ǂ̔����قǂ̒����B4���u�ǂ�䰕Ǐ㉏�̓����ɂ킸���ɔ��B������x�B䰕ǂ̌����⍂���́A�̂�Q�̂ɂ���č����傫���B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�A�ʒr�̊�Տ�ȂǂŌ�����B
�����ł̕��z�F��t���َR�E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{��͂���܂�Coelastrea aspera �p���J���m�R�L�N���C�V�i�{�K�C�h���f�ڎ�j��Favites pentagona �S�J�N�L�N���C�V�Ȃǂƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
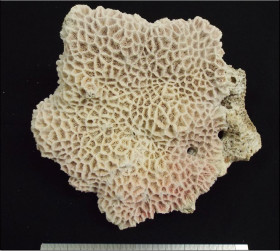
|
|
| |
Coelastrea sp. 2
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Coelastrea Verrill, 1866 �p���J���m�R�L�N���C�V��(�V��) |
|

�B�e�F�[���T�L(���V�\�s�匴�A���[8m)

|

䰂̒��a��8mm�`1.2cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�D�ΐF�`���F�B�אڌ̂Ƃ̋��E�����ĂŁA���ƌ��Ղ͏�ɔ����ۂ��B���Ղ́A���S�̌����͂ނ悤�Ɋ�ɐ���オ�邱�Ƃ������B
���i�̓����F�̂͂��ۂ݂�тт����p�`�ŁA�o��r���͍̌̂ג����L�т����p�`�Ɍ�����B�����̊O�ςƂ͈قȂ�A�̔z��͊��S�ȃZ���I�C�h�^�ŁA�o��r���ł͂Ȃ��̂�䰂̒��a��8mm�`1.2cm�قǂɂȂ�B�u�ǂ�4���܂Ŕ��B���A䰌a��1cm�ɖ����Ȃ����̂�40���O��B������䰕Ǐ㉏����킸���ɓ����ɒ���o�������ƁA䰕Ǔ����ɉ����ċ}�X�ŗ������ށB�u�ǂ̏㉏�ɂ́A��[�ɕ����̔����ˋN���������������悭���B����B�u�ǂ̌�����䰕Ǐ㉏�ł̓ˏo�x���́A�����ɂ��Ⴂ�͂قƂ�ǂȂ���l�Ɍ�����B�������̂ɂ���ẮA1���u�ǂ����̊u�ǂ���䰓��ɂ�蒣��o�����Ƃ�����B1���E2���u�ǂ͎����܂ŒB����B�����̊u�ǂ̓����ɂ̓p����t���`������邱�Ƃ����邪�A�s���ĂŖڗ����Ȃ��B3���u�ǂ͎����̎�O�܂ŁA4���u�ǂ͂���ɂ��̎�O�܂Řp�Ȃ��Ȃ���L�сA��u�ǂɗZ������B�����͒��a2�`5mm�قǂ̑ȉ~�`�ł悭�ڗ����A1���E2���u�ǂ̓�������L�т������̃g���x�L�����Ō`������Ă��邱�Ƃ��悭������B䰕ǂ��k���Ō��ŁA�����͊����㉏�܂œ�����1mm�قǁB
�������F�J���I�ȏʎΖʂ̐��[10m�Ȑ[�Ō�����B�J���I�Șp���ł́A�����[5m�ł������邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{��́A�����̓�������Dipsastraea �L�N���C�V���AFavites �J���m�R�L�N���C�V����Oulophyllia �I�I�i�K���T���S���̎�ƍ�������Ă���\��������B����č����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Cyphastrea confesta Nemenzo, 1959
�}�_���g�Q�L�N���V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Cyphastrea Milne Edwards and Haime, 1848 �g�Q�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[6m)

|

䰌a��1�`3mm�B
|
|
�����`�F�핢��`�����Q�̂ŁA�Q�̕\�ʂ��s�K���ɐ���オ��B��Ղ��L�������Ȃ���A���a50cm�قǂ̑�Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F�F�ʂ͎�ɉ��F�`�D���F�Ȃǂ̖��邢���̂��������A�H�ɈÊ��F�̂��̂�������B�Q�̕\�ʂŐ���オ�����Ƃ���͐F�������A�E�Ƃ���͔Z���Ȃ�A�S�̓I�ɂ܂���͗l�Ɍ����邱�Ƃ������B1���E2���u�ǂ��ˏo���Ă���̂������ł��悭�킩��B
���i�̓����F�͉̂~�`�ŁA䰌a��1�`3mm�A�傫����Ԋu�͕s�����ɂȂ�₷���B1���E2���u�ǂ́A䰕Ǐ㉏�Ŕ�����Ă悭�ˏo���A䰂̒��S�Ɍ������ċ}�X�ŗ������ށB2���܂ł̊u�ǐ���10���̂Ƃ�������B3���u�ǂ͔��ɔ����Z���ڗ����Ȃ��B�]�́A�ˏo�����̂������Ζő��ɔ��B���Ȃ��B�����̓�������A䰕ǂ����܂�ˏo���Ȃ����ɂ́A�����̌̂��~����ɓˏo�����悤�Ɍ�����B�̊Ԋu���L���J���������ł́A�����\�ʂɑ傫���̑�����������ˋN���悭���B����B�܂������ɂ́A���їނ̐��ǂƎv���錊��a���s�K���ɔ��B���邪�A�����ŔF�����邱�Ƃ͍���B
�������F�ʒr���ʌ���ʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Cyphastrea microphthalma �g�Q�L�N���C�V��C. ocellina �q���g�Q�L�N���C�V�i����Ƃ��ɖ{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B�a���͐���(2013)�Ɋ�Â��B
�����EVeron (1995)�ł̏o���L�^�F���iFavia favus�Ƃ��ċL�^�j

|
|
| |
Cyphastrea japonica Yabe and Sugiyama, 1932
�j�z���g�Q�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Cyphastrea Milne Edwards and Haime, 1848 �g�Q�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[5m)

|

䰌a��1�`2.5mm�B
|
|
�����`�F�핢��`���Q�̂ŁA�Q�̕\�ʂɃR�u��`�~����̓ˏo�����`������邱�Ƃ�����B�Օ��I�Ȋ��ł͒��a30cm����傫�ȉ��Q�̂ɂȂ邱�Ƃ�����B
��̕��̐F�ʂƓ����F�F�ʂ͉��ΐF�`�W���F�B�̂̓h�[����ɐ���オ���Ă���悤�Ɍ����A�̊Ԃɂ͑��їނ̐��ǂƎv���鏬���ȉ��ˏ�ˋN���F�߂��邱�Ƃ�����B
���i�̓����F�͉̂~�`��䰌a��1�`2.5mm�Ə����߁A�傫����Ԋu�͂悭�����B1���E2���u�ǂ̒����E�����͂قړ����œˏo�����A䰐S�Ɍ������Ċɂ₩�ɌX����B�u�Ǐ㉏�ɋ������悭���B����ꍇ�́A�u�Ǐ㕔���ˏo����䰓����h�[����ɐ���オ���Č�����B2���܂ł̊u�ǐ���10���̂Ƃ�������B3���u�ǂ͖ڗ����Ȃ��B�]���������A�u�ǂ̉�������������ˋN�����сA�]��Ɍ����邱�Ƃ�����B�����\�ʂɂ͑傫���̂悭������������ˋN�����B���A�̂����������߂��悭�ڗ��B
�������F�Օ��I�Ȋ�ʎΖʂ̐��[10m�Ȑ��A���D���͐ς�����p�̐��[5m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�a�̎R���V�����[�`��q���B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͍��m���y�������s�O��B�{���Cyphastrea microphthalma �g�Q�L�N���C�V�i�{�K�C�h���f�ڎ�j��C. serailia �t�J�g�Q�L�N���C�V�Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
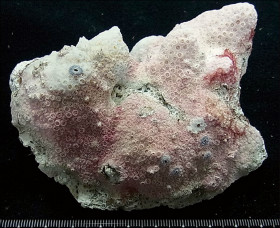
|
|
| |
Cyphastrea serailia (Forskål, 1775)
�t�J�g�Q�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Cyphastrea Milne Edwards and Haime, 1848 �g�Q�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[6m)

|

䰌a��1.5�`3.5mm�B
|
|
�����`�F�핢��`���Q�̂ŁA30�`40cm�قǂ̔�����ɂȂ邱�Ƃ����Ȃ��Ȃ��B�ʒr���ʌł́A�T���S�I��핢���Ĕ�Œ����̋���Q�̂ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F��ɒW���F�`���F�B�����́A�̗̂֊s���`�̂悭�����������Ȋ�(�h�[�i�c��)�Ɍ�����B���Ԃł��G���L���Ă��邱�Ƃ�����B
���i�̓����F�͉̂~�`��䰌a��1.5�`3.5mm�A�傫��������A�Ԋu�͌Q�̂��Ƃɂ悭�����B�܂��A�̊Ԋu���L���ƌ̒��a���傫���A�����Ə������Ȃ�X���ɂ���B�u�ǂƘ]��3���܂ł悭���B����B1���E2���u�ǂ́A䰂̒��S�Ɍ������Ċɂ₩�ɌX����B3���u�ǂ�1���E2���u�ǂɔ�ׂĔ����Z�����A䰕Ǐ㉏�ł͌����⍂���������x�ɂȂ�B��������ɔ�ׁA�{���䰕ǂ��悭���B����B�����̓�������A�u�ǂ�]���ˏo���Ȃ����ɂ́A�̂����~����ɓˏo���₷���B�����\�ʂ�������ˋN���悭���B���邪�A�ׂ��Z�����ߖڗ����Ȃ��B����ɁA�A����ɂȂ��������\�ʂ��悭�ڗ��B
�������F�ʒr�E��ʌ̐`�ʎΖʁA���Օ��I�Ȋ�ʎΖʁ`�Օ��I�ȓ��p�ōL�͈͂Ɍ�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Cyphastrea chalcidicum �R�g�Q�L�N���C�V�i�{�K�C�h���f�ڎ�j��C. japonica �j�z���g�Q�L�N���C�V�Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FFavia lizardensis�Ƃ��ċL�^�����\������

|
|
| |
Dipsastraea favus (Forskål, 1775)
�i�~�L�N���C�V�i���́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��Óc�A���[10m)

|

䰂̒��a��8mm�`1.5cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F���F�`�Ԋ��F�B���Ղ͍L��������B
���i�̓����F�͉̂~�`�`���тȑȉ~�`�A䰂̒��a��8mm�`1.5cm�A�Q�̂��ƂɌ̂̑傫���͂悭�����B�̔z��̓v���R�C�h�^�����A�̊Ԋu�������Ȃ�₷���A�Q�̒����ł͈ꕔ�Z���I�C�h�^�ɂȂ邱�Ƃ�����B䰕ǂ͔��߂ŁA�ˏo�����肷�ڂ肷�邱�Ƃ͂قƂ�ǂȂ��B�u�ǁE�]��4���܂Ŕ��B���邪�A3���E4���u�ǂ��悭���B����ꍇ�Ƃ��Ȃ��ꍇ������B4���u�ǂ܂ł悭���B�����̂ł́A1���E2���u�ǂ͎����܂ŁA3���u�ǂ������̂�����O�܂ł悭�L�т�B���̂Ƃ�4���u�ǂ́A䰕Ǐ㉏�ł͂悭���B���邪�A䰓��ł�䰕ǂɉ����Ă킸���ɔ��B������x�B�u�ǁE�]��䰕Ǐ㉏�œˏo���Ȃ��B1�`3���u�ǂ́A䰕ǂ̓����ɉ����ċ}�X�ŗ������ނ��A�p����t�̔��B�ƂƂ��ɓr�����玲�����ɒ���o���B�]�͒Z���A�̊Ԃŋ��L���邱�Ƃ������B�u�ǁE�]�̏㉏�ɂ͋������悭���B�A���Ԋu�Ŕz��B�܂������Ɗu�ǁE�]�̑��ʂɂ́A��[�̐����������ˋN���悭���B����B���̂��ߍ��i�\�ʂ͂�����Č����邱�Ƃ������B�����͉~�`�`�ȉ~�`�ŁA���a��1.5�`2mm�B
�������F�ʒr�E��ʌ̐�A���Օ��I�ȏʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F���c�E���c(1989)�ŃX�{�~�L�N���C�V�̘a�����^����ꂽ���Dipsastrea speciosa �L�N���C�V�̉\���������B�܂��AD. favus �̃^�C�v�W�{��䰕ǂ��ˏo�����A�̂����ڂ�Ō����邱�Ƃ͂Ȃ��B����Ė{��̘a���́A����E����(1985)�������i�~�L�N���C�V�ɉ��̂���B�{���D. speciosa �L�N���C�V�̂ق��AD. cf.favus ��Favites virens �I�I�J���m�R�L�N���C�V�ƍ�������Ă���B����āA�{��̍����ł̐������Č�������K�v������B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FFavia danae�Ƃ��ċL�^�����\������
|
|
| |
Dipsastraea (Forskål, 1775)
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��8mm�`1.5cm�B
|
|
�����`�F��ɉ��Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�ŁA�Q�̑S�̂��D�F�����������̂�A�Q�̕\�ʂ��s�K���Ȃ܂���͗l�ɂȂ邱�Ƃ�����B�̊Ԋu�͍L�������A䰕Ǐ㕔�͂��ꂢ�ȃh�[�i�c��Ɍ�����B
���i�̓����F�͉̂~�`�`���тȑȉ~�`�ŁA䰂̒��a��8mm�`1.5cm�قǂŁA1cm�O��̂��̂������B�̔z��͊��S�ȃv���R�C�h�^�ŁA��l�ɕ��z����B䰕ǂ͌��߂ŁA��₷�ڂ݂Ȃ���1mm�قǓˏo����B�u�ǁE�]��4���܂ł悭���B���A䰕Ǐ㉏�Ŕ�����A1mm���x�ˏo����B1���E2���u�ǂ͎����܂ŒB����B3���u�ǂ�1���E2���u�ǂ�2/3�`1/2�̒����ŁA�ꕔ�͎����܂ŐL�т���̂�����B4���u�ǂ͒����Ă�1���E2���u�ǂ�1/2�����B�]�͎����ɊW�Ȃ����������ŁA�אڌ̂Ƃ̋��E�܂Ŕ��B����B�����܂ł܂��͂��̋߂��܂ŐL�т�1�`3���u�ǂ̓����ɂ́A�������͂ނ悤�ɂ���������p����t�����B����B�u�ǂ́A䰕Ǐ㉏����p����t�܂ŋ}�X�ŗ������ނ̂ŁA䰂͍L���[��������B�u�ǁE�]�̏㉏�ɂ́A��[�̂悭��������������R�ƕ��ԁB���ʂɂ�������ˋN�����B���邪�������A����ł͕�����ɂ����B�����͒��a��2�`3mm�̉~�`�`�ȉ~�`�ŁA�悭�ڗ��B
�������F���Օ��I�Ȋ�ʎΖʂ̐��[15m�Ȑ�Ō�����B�J���I�Șp�ł͂��ł�������B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���ȓ�`��q���B��q���ł͕��ʎ�B
�⑫�F�{���Dipsastraea favus �i�~�L�N���C�V��D. speciosa �L�N���C�V�ƍ�������Ă���A��q���Ȗk�ł̐������Č�������K�v������B
* ��q�����L�^��

|
|
| |
Dipsastraea (Veron, Pichon and Wijsman-Best, 1977)
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��5mm�`1.2cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Q�̕\�ʂ��܂���͗l�ɂȂ�A���X�s�K���ɊD�F�≩�F�ɂȂ邱�Ƃ������B
���i�̓����F�̂͂��ꂢ�ȉ~�`�`���ȉ~�`�ŁA䰂̒��a��5mm�`1.2cm�ŁA������7�`9mm�B�̔z��͊��S�ȃv���R�C�h�^�ŁA��l�ɕ��z���邪�A�̊Ԋu�͋������̂�����L�����̂�����B䰕ǂ͌����A1�`3mm�قlj~����ɓˏo����B�u�ǁE�]��4���܂Ŕ��B���A���Ԋu�ł��ꂢ�ȕ��ˏ�ɔz��B1�`3���u�ǂ͎����ɒB����B4���u�ǂ͑��̊u�ǂ̔������x�̒����ŁA�����܂ŒB���Ȃ��B�u�ǂ�䰐S�Ɍ������ċ}�X�ŗ������ނ��A1���E2���u�ǂ̓����ɔ��B����p����t�ɂ���āA䰂͂���قǐ[�������Ȃ��B�]�͊u�ǂ��������A�����ɂ�炸�قړ��������ŁA�ˏo�̂������ΒZ���B�u�ǁE�]�̏㉏�ɂ́A�����������قړ��Ԋu�Ŕz��B�܂��u�ǁE�]�̑��ʂɂ́A��[�̉s��������Z��������ˋN���悭���B����B�����͒��a1.5�`2mm�قǂ̉~�`�ł悭���B����B
�������F���Օ��I�Ȋ�ʎΖʂ̐��[15m�Ȑ�Ō�����B�J���I�Șp���ł͂��ł�������B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Dipsastraea lizardensis ���U�[�h�L�N���C�V�i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F���iFavia pallida�Ƃ��ċL�^�j
|
|
| |
Dipsastraea aff. maritima Verrill, 1872
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��7mm�`1.2cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F�W�ΐF�≩�ΐF�`�W���F�B���Ղ͖��邢�F�ɂȂ�B�̂����ڂ݂Ȃ������ɓˏo�����l�q���悭������B
���i�̓����F�͉̂~�`�`�ȉ~�`�A䰂̒��a��7mm�`1.2cm�قǁB����Q�̏�ł�䰌a�͕s�����ŁA�̔z��͌̊Ԋu���s�K���ȃv���R�C�h�^�B�u�ǁE�]��4���܂Ŕ��B�B1���E2���u�ǂ͂������A�����܂ŒB����B1���E2���u�ǂɔ�ׂ��3���u�ǂ͂킸���ɒZ���A4���u�ǂ�1/3�`�����̒��������Ȃ��B�]�͂悭���B���A�����͊u�ǂƓ����x������ȏ�ɒB����B���̂��ߌ̂��悭���ڂ�Ō�����B�u�ǁE�]�̏㉏�ɂ́A��[�̐�����������悭���B���A�قړ��Ԋu�Ŕz��B���̂��߁A�悭�L�т��]�ł͂��̏㉏���_����Ɍ����₷���B�u�ǁE�]�⋘���̕\�ʂɂ́A������������̑������悭���B����B�����͉~�`�`�ȉ~�`�Œ��a��1.5�`2mm�ɂȂ邪�A���B�������̂▢���B�̌̂�������B
�������F���Օ��I�ȏʎΖʂ��ʎΖʂŌ�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�{���Dipsastraea maritima �i�{�K�C�h���f�ڎ�j�̂ق��AD. favus �i�~�L�N���C�V�AD. speciosa �L�N���C�V��D. danai �A�U�~�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă��邽�߁A�����ł̐����̍Č������K�v�B
* ��q�����L�^��
|
|
| |
Dipsastraea matthaii Vaughan, 1918
�A���L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��6mm�`1.2cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F�D�F���������ΐF�܂��͊��F�B䰓������F�Ȃǂ̖��邢�F��Z�����F�ł܂���͗l�ɂȂ邱�Ƃ������B䰓��͐��Ղ͍L��������B
���i�̓����F�̂͂��ꂢ�ȉ~�`�`���ȉ~�`�ŁA�Q�̒����̌̊Ԋu�������Ƃ���ł͂��p�����ȉ~�`�ɂȂ邱�Ƃ�����B䰂̒��a��6mm�`1.2cm�قǂŁA8mm�`1cm�̂��̂������B䰕ǂ͔����ˏo���Ȃ��B�u�ǁE�]��4���܂Ŕ��B�A䰕Ǐ㉏�Ŕ������B1���E2���u�ǂ͒����A�����܂ŒB����B�����̌X�͊ɂ₩�ŁA�����ɂ̓p����t�����B���邽�߁A䰓��͔��ɐ�����B3���u�ǂ͎����̂�����O�܂ŐL�т邱�Ƃ����邪�A������1���E2���u�ǂ�2/3�`1/2�̒����B4���u�ǂ͒����Ă�1���E2���u�ǂ�1/2�����B�u�ǁE�]�̏㉏�ɂ͋������悭���B���A�u�ǁE�]�Ƌ����̑��ʂɂ͔�����������ˋN���悭���B����B���̂��߁A���i�\�ʂ�������Č����₷���B�����͖��ĂŁA���a��1.5�`2mm�̉~�`�܂��͑ȉ~�`��悷��B�]�͔��B���邪�Z�߂ŁA�����L�т邱�Ƃ͋H�B
�������F�ʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō�����B�ʒr���ʌł����܂Ɍ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Dipsastraea pallida �E�X�`���L�N���C�V���͂���D. speciosa �L�N���C�V��D. lizardensis ���U�[�h�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă��邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F���iFavia speciosa�Ƃ��ċL�^�j
|
|
| |
Dipsastraea pallida (Dana, 1846)
�E�X�`���L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��6mm�`1cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�B��F��W���F�A�Q�̕\�ʂ��܂���͗l�ɂȂ������̂�������B���Ղ͏������A���̕����̐F�ʂ����邭�悭�ڗ��B
���i�̓����F�̂͂��ꂢ�ȉ~�`�`���ȉ~�`�ŁA䰂̒��a��6mm�`1cm�B�̔z��͊��S�ȃv���R�C�h�^�ŁA䰕ǂ͔����A1�`3mm�قlj~����ɓˏo����B�ˏo�̒��x�͌Q�̂��Ƃɂ悭�����B�u�ǁE�]�͔����A4���܂Ŕ��B���A䰕Ǐ㉏�ł悭�ˏo����B1���E2���u�ǂ͎����܂ŒB���A�����Ƀp����t�����B����B���3���u�ǂ�1���E2���u�ǂ�2/3�`1/2�A4���u�ǂ�1/3�̒����B�]�͂悭���B���A�̊Ԋu���L���Ƃ���ł͑Ή�����u�ǂ��������L�тĂ悭�ڗ��B�u�ǂƘ]�̏㉏�ɂ͕����������悭���B����B�܂����ʂɂ́A������������ˋN�����B����B�����͉~�`�`�ȉ~�`�Ŗ��āA���a��1.5�`2mm�قǁB䰕ǂ�䰕Ǐ㉏�̊u�ǁE�]���ˏo���銄�ɂ́A���B�����p����t�Ǝ����̑��݂ɂ���āA䰓��͂���قǐ[�������Ȃ��B
�������F�ʒr���ʌ̐���ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B��ʎΖʂ�Օ��I�ȓ��p�ł����܂Ɍ�����B
�����ł̕��z�F�a�̎R�����{�E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Dipsastraea speciosa �L�N���C�V���͂���D. matthaii �A���L�N���C�V��D. lizardensis ���U�[�h�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���B����č����ł̐������Č�������K�v������B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FFavia veroni�Ƃ��ċL�^�����\������
|
|
| |
Dipsastraea cf. pallida
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[10m)

|

䰂̒��a��5mm�`1cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�̂��悭�ˏo���A�̊Ԋu���L���̂Ř]�̕������悭�ڗ��B
���i�̓����F�̂͂��ꂢ�ȉ~�`�`���ȉ~�`�ŁA䰂̒��a��5mm�`1cm�قǁB�Q�̏�ł̌̔z��͊��S�ȃv���R�C�h�^�ŁA�̊Ԋu�͍L���A��l�ɕ��z����B䰕ǂ͌����A�~�����2�`5mm�قǓˏo����B�u�ǁE�]��4���܂Ŕ��B�A䰕Ǐ㉏�Ŕ�����A1mm���x�ˏo����B�u�ǂ�䰕Ǐ㉏���玲���Ɍ������ċ}�X�ŗ������ނ��߁A䰓��͍L��������B1���E2���u�ǂ�3���u�ǂ̈ꕔ�͎����܂ŒB����B�c���3���u�ǂ̒����́A1���E2���u�ǂ�1/2�`2/3�̒��x�A4���u�ǂ�1/2�����B1���E2���u�ǂ̓����́A�킸���ɔ�����ăp����t���`������邱�Ƃ�����B�]�͎����ɊW�Ȃ��ˏo����䰕ǂ̊O������̋��E�܂ł悭���B����B�u�ǁE�]�̏㉏�ɂ͐�[�̐�����������K�����������B����B�܂��A�u�ǁE�]�E�����̑��ʂɂ�������ˋN�����B����B�����͒��a��2�`3mm�̉~�`�`�ȉ~�`�B
�������F���Օ��I�Ȋ�ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�`��q���B��q���ł͋H�B
�⑫�F�{���Dipsastraea pallida �E�X�`���L�N���C�V�ƍ�������Ă���\�������邽�߁A�����ł̏ڍׂȐ����̒������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FE. lamellosa�Ƃ��ċL�^�����\������
|
|
| |
Dipsastraea speciosa (Dana, 1846)
�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[4m)

|

䰂̒��a��5mm�`1.5cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B���Ղ̐F�����ƈقȂ邱�Ƃ������B䰂͍L���[��������B
���i�̓����F�͉̂~�`�`�ȉ~�`�ŁA䰂̒��a��5mm�`1.5cm�A8mm�`1cm�̌̂������B䰕ǂ͂������A����Ɍ������Ă킸���ɂ��ڂ݂Ȃ���1�`3mm�قǓˏo����B�̔z��̓v���R�C�h�^�����A�̊Ԋu�������Ȃ�₷���A�Q�̒����ł͈ꕔ�Z���I�C�h�^�ɂȂ邱�Ƃ�����B�u�ǁE�]�͔����A4���܂Ŕ��B����B������3���E4���u�ǂ̒������s�����ŁA�z����Č�����B1���E2���u�ǂ�3���u�ǂ̈ꕔ�͎����܂ŒB����B�c���3���u�ǂ̒����́A1���E2���u�ǂ�1/2�`1/3�̒��x�A4���u�ǂ�1/3�����B�u�ǂ�䰕Ǐ㉏������Ɍ������Ă��܂�ˏo�����A��������䰐S�Ɍ������ċ}�X�ŗ������ށB�܂��A1���E2���u�ǂ̓����ł̃p����t�̔��B�������B���̂��߁A䰂͍L���[��������B�]��䰕ǂ̊O���ʂł͂悭���B���邪�A������܂Ŕ��B���A�̊ԂŘ]���A�����邱�Ƃ͂قƂ�ǂȂ��B�u�ǁE�]�̏㉏�ɂ͋������A���ʂɂ�������ˋN�����ꂼ�ꔭ�B���邪�A�Ƃ��ɏ������Ėڗ����Ȃ��B�����͉~�`�`�ȉ~�`�A���a��1.5�`2mm�B
�������F�ʎΖʂ̐[�ꂩ��ʒr���ʌA���p�`���Օ��I�Ȋ�ʎΖʂōL��������B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Dipsastraea favus �i�~�L�N���C�V��D. pallida �E�X�`���L�N���C�V���͂��ߑ����̓�������ƍ�������Ă���B
* ��q�����L�^��

|
|
| |
Dipsastraea aff. veroni (Moll and Borel-Best, 1984)
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Dipsastraea de Blainville, 1830 �L�N���C�V�� |
|

�B�e�F�[���T�L(���V�\�s��Óc�A���[22m)

|

䰂̒��a��1.5cm�ȏ�B
|
|
�����`�F��ɉ��Q�́B
��̕��̐F�ʂƓ����F�D���F�`�D�ΐF�ŁA���Ղ͂��W���F�ɂȂ邱�Ƃ��قƂ�ǁB
���i�̓����F�̂͂��тȂ܂��͊p�����ȉ~�`�A���a��1.5cm�ȏ�Ƒ傫���A3cm�߂��ɂȂ���̂�����B�̂̓v���R�C�h�^�ɔz�A�̊Ԋu��2mm���x�����Ȃ����̂���5mm�ȏ㗣��Ă�����̂܂ŗl�X�B䰕ǂ͔����A�����\�ʂ���قƂ�Ǔˏo���Ȃ��B�u�ǁE�]�͔����A4���܂Ŕ��B���A䰕Ǐ㉏�ł悭�ˏo����B�u�ǂ�䰐S�Ɍ������ċ}�X�ŗ������ށB���̂���䰓���5mm�`1.2cm�قǂ̐[���ɂȂ�B���B�r����4���u�ǂ������A�u�ǂ͑S�Ď����܂ŒB����B�u�Ǔ����ł̃p����t�̔��B�͈����B�]�́A�̊Ԋu�����ȌQ�̂ł͒Z�����A�̊Ԋu���L�����̂ł͌̓��m��A������悤�ɂ悭���B����B�u�ǂƘ]�̏㉏�ɂ́A��[�����A�傫���̑��������������Ԋu�Ŕz��B�܂��u�ǁE�]�E�����̑��ʂɂ́A��[��������ˋN�������ɑ��Đ����ɕ��ԁB�����͉~�`�`�ȉ~�`�ł悭���B���A���a��2�`6mm�قǁB��������ɔ�ׂČQ�̗��ʂł̃G�s�e�J�̔��B���ɂ߂Ĉ����B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō����A����Ȑ�Ō����邱�Ƃ͋H�B
�����ł̕��z�F�a�̎R�����{�E���茧�ܓ����]���`����������E���B��q���ł͋H�B
�⑫�F�{��͌̒��a���ގ�����Dipsastraea veroni �A�o���L�N���C�V��D. maxima �E���g���L�N���C�V�i����Ƃ��ɖ{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Echinopora cf. gemmacea (Lamarck, 1816)
�I�I�����E�L���E�L�b�J�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Echinopora Lamarck, 1816 �����E�L���E�L�b�J�T���S�� |
|

�B�e�F������^(���V�\�s��Óc�A���[10m)

|

䰌a��3�`5mm�B
|
|
�����`�F�핢��Q�́B�������A�Q�̗��ʂ����S�Ɋ�ՂɌŒ����邱�Ƃ͂Ȃ��B�Q�̎����͗t��ɂȂ��Ē���o�����Ƃ������B
��̕��̐F�ʂƓ����F�����F�A�N���[���F��ΐF�`���F�B�Q�̎����ŐF���W���Ȃ邱�Ƃ������B�Q�̕\�ʂ̓U���U�����Č�����B
���i�̓����F�̂͒������������`�`�~���`�ŁA䰌a3�`5mm�A����Q�̏�ł̑傫���A������ˏo�̒��x�͕s�����B�u�ǂ�3���܂Ŕ��B���A���܂ɕs���S��4���u�ǂ������邱�Ƃ�����B1���E2���u�ǂ͎����܂ŒB����B1���u�ǂ́A2���u�ǂɔ�ׂ�䰕Ǐ㉏�߂��ł������Ȃ邪�A���̈Ⴂ�͓���ŋ�ʂ��ɂ����B3���u�ǂ͔����A䰕Ǐ㉏����䰓��Ɍ������Ă킸���ɐL��������x�����A�����߂��܂ŐL�����Ă�����̂�����B1���E2���u�ǂ̏㉏�⋤���\�ʂł́A�����⑾���̂悭�����������Ɛj��ˋN�����ɔ��B����B���ĂȘ]�͔��B���Ȃ����A�j��ˋN��䰕ǂ̊O����Q�̎����ŋK������������Ř]��Ɍ�����B1���u�Ǐ㉏�̋����̐��́A�p����t�����܂߂��4�����A�p����t�������B�܂��͒��Ԃ̋���2���Z�����Ă��鎞��2�`3�Ɍ�����B2���u�ǂł�䰕Ǐ㕔�t�߂�2�m�F�ł���B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B�ʒr���ʌł������邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͋H�B
�⑫�F�{��̌̂̑傫���A�u�ǁE�]�̏㉏�̋����̌`���䰕ǂ̓ˏo�x���́AEchinopora gemmacea �i�{�K�C�h���f�ڎ�j�̃^�C�v�W�{�̂����ƈقȂ��Ă���B�܂��{���E. lamellosa �����E�L���E�L�b�J�T���S�Ƃ���������Ă���B����Ė{��́A�X�Ȃ镪�ފw�I�����ƍ����ł̐����̍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Favites halicora (Ehrenberg, 1834)
�}���J���m�R�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[4m)

|

䰂̒��a��8mm�`1.2cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B���Ղ܂��͊u�ǁE�]�̏㉏������߂̐F�ʂɂȂ邱�Ƃ�����B䰕ǂ͌����A䰂͐A���Ղ͏�����������B�P���u�ǂ��s�K���ɓˏo�����̂������邱�Ƃ�����B
���i�̓����F�͉̂~�`�`���ۂ݂�тт����p�`�ŁA䰂̒��a��8mm�`1.2cm�قǁB�����̔z��̓v���R�C�h�^�ɂ��Z���I�C�h�^�ɂ��Ȃ�B䰕ǂ�2mm�قǂŁA�����͊ۂ݂�ттĂ��邱�Ƃ������B�u�ǁE�]��4���܂Ŕ��B���A䰕Ǐ㉏�œˏo���邱�ƂȂ��A䰐S�Ɍ������ċ}�X�ŗ������ށB�������A�傫�Ȏ�����䰓��̐Ƃ���ł悭���B���邽�߁A䰓��͂���قǐ[�������Ȃ��B�P���E�Q���u�ǂ͊��S�Ɏ����ɒB���A�����̓����͂悭������āA�p����t�܂��̓p�������B���邱�Ƃ����Ȃ��Ȃ��B3���u�ǂ́A�����͂P���E�Q���u�ǂƂقړ��������A�������傫�����B�����̂������A����炪�����ɒB���邱�Ƃ͂Ȃ��B�S���u�ǂ͑��̊u�ǂɔ�ׂĔ����A䰕Ǐ㉏�ł킸���Ɍ`���������x�B�]�͒Z���A�قڒ����������Ă���B�u�ǁE�]�̏㉏�ɂ́A�ۂ��Đ�[�����A���ʂ�������ˋN��������������悭���B����B���̂��߁A�̂̕\�ʂ̓U�����Č�����B�����͉~�`�`�ȉ~�`�ŁA���a�͏�����1mm�A�傫�����̂�3mm�قǂɂȂ�B�����́A�悭�ˏo�����p����t��p���ň͂܂�邽�߁A�悭�ڗ��B
�������F�ʒr�E��ʌ̊�Տォ��ʎΖʂ��ʎΖʂ̐[��܂ōL��������B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Favites abdita �J���m�R�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ƃ悭��������Ă��邽�߁A�����ł̕��z�̍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F���iFavia rotundata�Ƃ��ċL�^�j
|
|
| |
Favites pentagona (Esper, 1795)
�S�J�N�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�i�c�r��(���V�\�s�Y�c�p�A���[6m)

|

䰂̒��a��6�`8mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂɒ��a����э�����1�`4cm�قǂ̉~����̓ˏo�����s�K���Ɍ`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɉ×ΐF�`�Ԋ��F�ŁA���ΐF��W���F�̂��̂�A䰕Ǐ㉏���������̂�A���Ղ̐F�ʂ����ƈقȂ���̂�������B�����ł��̂͂��ꂢ�ȑ��p�`�Ɍ����A�u�ǂ��K���������z�Ă���l�q���悭������B䰓��͐A���Ղ͏�����������B
���i�̓����F�̂͑��p�`�ŁA䰂̒��a��6�`8mm���قƂ�ǂŁA�`��͂悭�����Č�����B�̔z��͊��S�ȃZ���I�C�h�^�B䰕ǂ͊��2�`3mm�ƌ������A�����ł�0.5mm�ȉ��Ɣ����A������Ɍ�����B�u�ǂ�4���܂Ŕ��B���A�������Ƃɒ���������͂悭�����A���ˏ�ɋK���������z��B�u�ǂ�䰕Ǐ㉏�œˏo�����A䰕ǂ̓����ʂɉ����悤�ɔ��B����B���̂���䰓��́A�ĕ����Ɍ�����B1���E2���u�ǂ͓�������������A���Ăȃp���܂��̓p����t���`�������B3���u�ǂ�䰕ǂ̓����ʂ���䰐S���܂ŐL�����邪�A�����܂ŒB���邱�Ƃ͂Ȃ��B4���u�ǂ͒Z���A䰕ǂ̓����ʏ�ɔ��B����̂݁B�u�Ǐ㉏�ɂ͐�[���悭���A���ʂ�������ˋN���������ȋ������т�����Ɣ��B����B������1mm�ȉ��ŏ��������A������͂ނ悤�ɂ悭���B�����p����p����t�̑��݂ɂ��F�����₷���B
�������F�ʒr�E��ʌ̊�Տ��A�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Coelastrea sp. 1��Favites aff.pentagona �V���t���S�J�N�L�N���C�V�Ȃǂƍ�������Ă��邽�߁A��q���Ȗk�ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��

|
|
| |
Favites aff. pentagona (Esper, 1795)
�V���t���S�J�N�L�N���C�V�i�V�́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F������^(����q���n���̊≮�A���[8m)

|

䰂̒��a��7�`9mm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F��ɊD�ΐF��Ê��F�B�u�Ǐ㉏�������Ȃ�A�Q�̕\��(����䰕Ǐ㉏)�ɑ����~�����悤�Ɍ����邱�Ƃ������B���ƌ��Ղ̐F�ʂ��قȂ邱�Ƃ�����B
���i�̓����F�̂͑��p�`�`�ג����L�т����p�`�ŁA䰂̒��a��6mm�`1.2cm�ŁA7�`9mm�̂��̂������B�̔z��͊��S�ȃZ���I�C�h�^�����A���їނ̐��ǂƎv����Ǐ�Ⓑ���a��̊Ԍ����A䰕ǂ�2������悤�ɕs�K���Ɍ`������邱�Ƃ�����B䰕ǂ͊��1�`1.5mm�قǂ����A����Ɍ������ɂꔖ���Ȃ�A�����ł̌�����0.5mm�����ɂȂ�B�u�ǂ�4���܂Ŕ��B���A�����͎������Ƃ̈Ⴂ���s���āA�X�͂悭�����Ċɂ₩�Ɍ�����B䰌a��8mm�قǂ̌̂ł́A�u�ǐ���25�`30�����x�B1���E2���u�ǂ͎����܂ŒB�A�����͔�����A�p����t��p���������ɔ��B����B3���u�ǂ�1���E2���u�ǂ�1/2�`2/3�قǂ̒����ŁA�����ɒB���Ȃ��B4���u�ǂ͔��ɒZ���A䰕Ǔ����ʂ���킸���ɒ���o�����x�B�u�Ǐ㉏�ɂ́A�`���s�����ŕ\�ʂɐ���������ˋN�����������B����B�u�ǂ̑��ʂɂ́A������������̑�����������B������1�`2mm�قǂŒʏ�͔F�����₷�����A�����Ȍ̂�p����t��p���̔��B�������̂ł͕s���ĂŖڗ����Ȃ��B
�������F��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�`�g�J�����V���B��q���ł͂��܂Ɍ�����B
�⑫�F�{��͂���܂�Favites pentagona �S�J�N�L�N���C�V��Coelastrea sp. 1 �Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Favites rotundata Veron, Pichon and Wijsman-Best, 1977
�A�c�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��Óc�A���[10m)

|

䰂̒��a��1.5�`2.5cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F��ɗΐF�B��͓̂����ŁA�̂͗֊s���ۂ݂�ттĂ���A���ꂢ�ȃh�[�i�c�^�Ɍ�����B䰕Ǐ㉏�͒W���D�ΐF�A���Ղ͊D�F�Ŗ��邭�����邱�Ƃ������B�u�Ǐ㉏�������A䰂͐�����B1���u�ǂ��s�K���ɓˏo�����̂������������邱�Ƃ�����B
���i�̓����F�͎̂�ɉ~�`�`�ȉ~�`�ŁA䰂̒��a��1.5�`2.5cm�ŁA1.8cm�O��̂��̂������B�̔z��͊�{�I�Ƀv���R�C�h�^�B�̂̑傫���͕s�����ŁA�̂̊Ԋu�A�����A�ˏo�x�����s�K���ɂȂ�₷���B䰕ǂ͌����A�u�ǁE�]��4���܂Ŕ��B����B1�`3���u�ǂ͌��߂ŁA䰕Ǐ㉏�ł͂���Ɍ����Ȃ�B�u�njX���͊ɂ₩�ŁA�㉏�ɂ͋������A�����ɂ̓p����t�����B���₷�����߁A䰓��͏�ɐB1���E2���u�ǂ͎����܂ŒB���邪�A3���u�ǂ͂�����2/3�قǂ̒����ɂ����Ȃ炸�A�����ɂ͒B���Ȃ��B4���u�ǂ͔����A䰕Ǐ㕔�ł킸���ɔ��B����݂̂����A�����H��3���u�ǂƓ����x�̒����܂ŐL�т邱�Ƃ�����B�]��䰕ǂ̂����O���ɒZ�����B���邪�A�̊Ԋu���L���Ƃ���ł͒����Ȃ邱�Ƃ�����B�u�ǁE�]�̏㉏�̋����͌����ŁA��[�����[�u��ɂȂ邩�A�܂��͂悭����Ă���A�`��傫����Ԋu�������Ă���B�����̑��ʂ��炻�̉����̊u�Ǒ��ʂɂ́A������������ˋN�����ɔz��B�����͉~�`�`�ȉ~�`�ł悭�ڗ����A���a��3�`4.5mm�قǂɂȂ�B
�������F�J���I�ȏʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�J���I�Șp�ł͂��ł�������B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{���Favites cf.rotundata ��F. aff.rotundata �ƍ�������Ă���\��������B���ɉ���{���Ȗk�ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F���iMontastrea valenciennesi�Ƃ��ċL�^�j
|
|
| |
Favites cf. rotundata Veron, Pichon and Wijsman-Best, 1977
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�[���T�L(����q���n���̊≮�A���[14m)
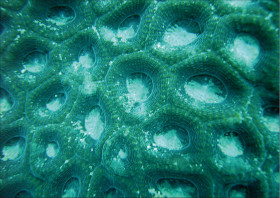
|

䰂̒��a��1.5�`2.5cm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂɒ��a�ƍ������ő��5cm���x�̃R�u��̓ˏo�����s�K���Ɍ`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�ŁA�Ԃ݂�тт����̂�����B��̂������ŁA䰂͐A�̂͊ۂ݂�ттĂ��邪�A�אڌ̂̋��E�͂��ꂢ�ȑ��p�`�Ɍ�����B�̂��ˏo���ėl�X�ȕ������������Ƃ͂Ȃ��B
���i�̓����F�̂͂��тȑ��p�`�ŁA�召�l�X�Ȍ̂��Z���I�C�h�^�`�v���R�C�h�^�ɔz��B䰂͒��a1.5�`2.5cm�قǂ����A�H��3cm�߂��̂������邱�Ƃ�����B䰕ǂ͌���1�`2mm�Ŋ���璸���܂œ��������B������1cm���邱�Ƃ����邪�A�u�ǂ̌X���ɂ₩�Ȃ���䰓��͐[�������Ȃ��B�u�ǂ�5���܂łŁA������䰕Ǐ㉏�ł̓ˏo�x���͎������ƂɈقȂ�B���̂��߁A䰕Ǐ㉏��������ʂ̕������e���s�����Ɍ�����B1�`4���u�ǂ͂قړ������������A5���u�ǂ͂�┖���B1���E2���u�ǂ͒����A䰕Ǐ㉏�ł悭�ˏo���A�����܂ŒB����B3���u�ǂ̒����͕s���������A������O�܂ŐL�тĒ�u�ǂɗZ�����邱�Ƃ�����B4���u�ǂ́A�L�тĂ���̂��̂�1/2�`2/3�قǂ̒����܂ŁB5���u�ǂ͔��ɒZ���A䰕ǂ̓����ʂɂ킸���Ɍ�����̂݁B1�`3���u�Ǔ����ɂ̓p����t���`������邱�Ƃ����邪�A�قƂ�ǂ͕s���ĂŖڗ����Ȃ��B�u�Ǐ㉏�ɂ́A��[��������A�܂��͊ۂ����[�u��ɂȂ����������悭���B����B������䰕Ǒ��ŒZ���ď��������A�������ł͒����đ傫���B�����͑ȉ~�`�Ŗ��āA���a��2�`6mm�B
�������F��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���`��q���B��q���ł͋H�B
�⑫�F�{���Favites rotundata �A�c�L�N���C�V��F. aff.rotundata �ƍ�������Ă���B����č����ł̐����͍Č������K�v�B�܂��A���i�`�Ԃ��A�c�L�N���C�V�Ɨގ����邽�߁A����̍X�Ȃ镪�ފw�I�������K�v�ł���B
* ��q�����L�^��
|
|
| |
Favites aff. rotundata Veron, Pichon and Wijsman-Best, 1977
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�i�c�r��(���V�\�s�匴�A���[6m)
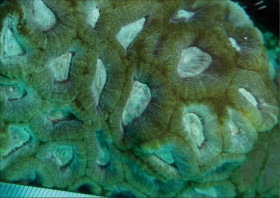
|

䰂̒��a��1�`2cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�B䰕Ǐ㉏�ƌ��Ղ͊D�F�������Ė��邭������B��͓̂��������A䰕ǂ������̗̂֊s���e�����߁A�̂̓h�[�i�c�^�Ɍ����ɂ����B䰂͐[���A���Ղ͍L��������B1���u�ǂ��s�K���ɓˏo���A���̏㉏�������ۂ��Ȃ邱�Ƃ�����B
���i�̓����F�͎̂�ɉ~�`�`�ȉ~�`�ŁA䰂̒��a��1�`2cm�ŁA����Q�̏�ł̂����傫���͕s�����Ɍ�����B�̔z��̓v���R�C�h�^�����A�̊Ԋu�������A�����I�ɃZ���I�C�h�^�Ɍ����邱�Ƃ�����B�̂̌�����ˏo�x���͕s�����Ɍ����邪�A�o�肵�ĊԂ��Ȃ������Ȍ̂ƁA�Q�̎����̌̂�䰕ǂ��ˏo����X��������B�u�ǂ�5���܂Ŕ��B���A5���u�ǂ������Ό����͂قړ����Ŕ����B������䰕Ǐ㉏�ł悭�ˏo���A䰐S�Ɍ������ċ}�X�ŗ������ށB���̂��߁A䰌a���킸䰓��͐[��������B�܂������ꕔ�̌̂ł́A1���u�ǂ������������Ȃ��āA�����ɒ���o�����Ƃ�����B1���E2���u�ǂ͎����܂ŒB���邪�A���̓����ł̃p����t�̔��B�͈����B3���u�ǂ͂����̓�����������O�܂ŁA4���u�ǂ�3���u�Ǔ����̂���Ɏ�O�܂ł����B���Ȃ��B�����̊u�Ǔ����́A��u�ǂ̂ق��ɘp�Ȃ��邽�߁A�u�ǂ̔z���ꂢ�ȕ��ˏ�Ɍ����Ȃ��B5���u�ǂ͔��ɒZ���A䰕Ǐ㉏�ł킸���ɔ��B����̂݁B�܂����̊u�ǂɔ�ׂĔ����A䰕Ǐ㉏�œˏo���Ȃ����߁A�ڗ����Ȃ��B�]�͓ˏo�����̂̊O���ʂł͖��āB�u�ǁE�]�̏㉏�̋����͌����ŁA��[�����[�u��ɂȂ邩�A�܂��͐���Ă���A�`��傫����Ԋu���悭�����B�����́A���a��3�`4mm�قǂ̉~�`�`�ȉ~�`�ŁA�悭���B����B
�������F�ʒr�E��ʌ̊�Տ��A�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���`����{���B��q���ł͋H�B
�⑫�F�{���Favites rotundata �A�c�L�N���C�V�ƍ�������Ă���\��������B����č����ł̐����͍Č������K�v�B�܂��{��̌`�Ԃ́ADipsastraea �L�N���C�V���Ƃ̗ގ������������߁A�X�Ȃ镪�ފw�I�������K�v�Ǝv����B
* ��q�����L�^��

|
|
| |
Favites valenciennesi (Milne Edwards and Haime, 1849)
�^�J�N�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�[���T�L(����q���剖���A���[2m)

|

�אڌ̂͘]�����L���Ȃ��B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F���F�`�D�ΐF�B䰕ǂ̓��������W���F�ɂȂ�A���Ղ��Â������邱�Ƃ������B�̂͂��p���Ă��邽�߁A�h�[�i�c��Ɍ����邱�Ƃ͏��Ȃ��A�Q�̕\�ʂ������Ɍ�����B
���i�̓����F�̂͊p�����~�`�`���p�`�ŁA䰂̒��a��5�`8mm�قǁB�̔z��͊��S�ȃv���R�C�h�^�ŁA�̊Ԋu�͋����B�אڌ̂̋��E�ɂ́A���їނ̐��ǂƎv����Ǐ�܂��͒Z���a��̊Ԍ����s�K���ɕ��z����B䰕ǂ͌������قƂ�Ǔˏo���Ȃ��B�u�ǁE�]�͌��߂ŁA䰕Ǐ㉏�ł͂���ɔ������B�����͒ʏ�4���܂Ŕ��B���邪�A4���̔��B�͈����B1���E2���u�ǂ͒����A䰕Ǐ㉏����}�X�ŗ�������A�����܂ŒB����B�����̓����͂�������Ă���A�p����t�����B���邱�Ƃ�����B3���u�ǂ�1���E2���u�ǂ�1/2�`2/3�̒����ŁA�������̂͂��̓�����1���E2���u�ǂ̑��ʂɗZ������B�]�̒����͎����Ɋւ�炸�����ŒZ���B�̊ԂŘ]�����L����ӏ��͂قƂ�ǂȂ��B�u�ǁE�]�̏㉏�̋����͐�[�̐�����~����ŁA����������Ȃ��琮�R�ƕ��ԁB�u�ǁE�]�E�����̕\�ʂɂ�������̑������F�߂��邪�����Ŗڗ����Ȃ��B�����͒��a1�`2mm�قǂ̉~�`�`�ȉ~�`�ł悭���B����B�Q�̍��i������ƁA�̋��E�ł��ꂢ�Ɋ���邱�Ƃ͏��Ȃ��B
�������F�ʒr�E��ʌ̊�Տ��A�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Favites aff.valenciennesi �j�Z�^�J�N�L�N���C�V�ƍ�������Ă�������(�[���E�쑺, 2009)�A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FFavites flexuosa�Ƃ��ċL�^�����\������
|
|
| |
Favites aff. valenciennesi (Milne Edwards and Haime, 1849)
�j�Z�^�J�N�L�N���C�V�i�V�́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

�אڌ̂������I�ɘ]�����L����B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B�Q�̕\�ʂ�ꕔ�̌̂�䰓����s�K���ɊD�F������A�܂���͗l�Ɍ�����B�X�̌̂̓L�N���C�V���̂悤�Ƀh�[�i�c�^�ɐ���オ���Č�����B
���i�̓����F�̂͊p�����~�`�`�ȉ~�`�A䰂̒��a��5mm�`1cm�ŁA7�`8mm�̂��̂������B�̊Ԋu�͈�l�ɋ������A�z��͊��S�ȃv���R�C�h�^�B�אڌ̂̋��E�ɂ́A���їނ̐��ǂƎv����Ǐ��a��̊Ԍ����e�̂��͂ނ悤�Ɉ�l�ɕ��z����B䰕ǂ͌����A��₷�ڂ݂Ȃ���1mm���x�ˏo����B�u�ǁE�]�͔������A4���܂Ŕ��B���A䰕Ǐ㉏�Ŕ�����A�킸���ɓˏo����B�u�ǂ̌X�͊ɂ₩�ŁA䰓��͐�����B1���E2���u�ǂ͎����܂ŒB����B3���u�ǂ��ꕔ�͂قڎ����܂ŒB���邪�A�c���1���E2���u�ǂ�1/2�`2/3�̒����ɂƂǂ܂�B4���u�ǂ�䰕Ǐ㉏�ł͂悭���B���邪�A䰓��ł�1���E2���u�ǂ�1/2�����̒����ɂ����Ȃ�Ȃ��B1�`3���u�ǂ̓����ɂ̓p����t�����B���₷���B�]��䰕ǂ̊O���ɉ����Ă悭���B����B�u�ǁE�]�̏㉏�ɂ͐��R�ƕ��������A���ʂɂ�������ˋN���悭���B����B�]�́A�אڂ���̊Ԃŏ㉏�̋������A������ӏ��������F�߂���B�����͂悭���B���A���a��1�`1.5mm�قǂ̉~�`�`�ȉ~�`�B�Q�̍��i������ƁA�̋��E�ł��ꂢ�Ɋ���₷���B
�������F�ʒr�E��ʌ̊�Տ��A�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{��͂���܂�Favites valenciennesi �^�J�N�L�N���C�V�ƍ�������Ă�������(�[���E�쑺, 2009)�A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Favites virens (Dana, 1846)
�I�I�J���m�R�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(����q���n���̊≮�A���[6m)

|

䰂̒��a��8mm�`1.6cm�B
|
|
�����`�F�����`���Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B�Q�̕\�ʂ�ꕔ�̌̂�䰓����s�K���ɉ��F�܂��͊D�F������A�܂���͗l�Ɍ����邱�Ƃ������B䰕ǂ������ˏo���Ȃ����߁A�̂��h�[�i�c�^�ɐ���オ���Č����邱�Ƃ͂Ȃ��B
���i�̓����F�̔z��̓v���R�C�h�^�`�Z���I�C�h�^�B�͉̂~�`�`���p�`�B䰂͒��a8mm�`1.6cm�ŁA���̒��Ԃ��炢�̌̂������B䰕ǂ͔����ēˏo���Ȃ��B�u�ǁE�]��5���܂Ŕ��B���A䰕Ǐ㉏�œˏo���Ȃ��B�v���R�C�h�^�Q�̂̏ꍇ�A䰕Ǐ㕔�����R�Ɍ�����B1�`4���u�ǂ͔����A䰕Ǐ㉏�łقړ��Ԋu�ɔz��B�u�ǂ̌X�͗l�X�����A�Q�̂��Ƃɂ͂悭�����B1���E2���u�ǂ͎����܂ŒB���A���̓����Ƀp����t�����B���邱�Ƃ�����B3���u�ǂ͎����̂���O�܂ŁA4���u�ǂ�3���u�Ǔ����̂���Ɏ�O�܂ł����L�тȂ��B�����̊u�Ǔ����́A��u�ǂ̂ق��ɘp�Ȃ��ėZ�����邱�Ƃ�����B5���u�ǂ͔��ɒZ���A䰕Ǐ㉏�Ƒ��ʂł킸���ɔ��B����B1�`4���]�͒Z���قړ��������ŁA1�`4���u�ǂɔ�ׂ�Ƃ������B�u�ǁE�]�̏㉏�ɂ͋������A�����̕\�ʂƊu�ǁE�]�̑��ʂɂ�������ˋN���悭���B����B�u�ǂ̋����̓��[�u��ŁA䰂̉����̂��̂قǓˏo����B�����́A���a��3�`4mm�قǂ̉~�`�`�ȉ~�`�B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ��ᏼ���ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Veron et al. (1977)��Favites flexuosa �i�{�K�C�h���f�ڎ�j�̃V�m�j���ɂ���Ă���B�������A�{��̊u�ǂ̌`��⎟�����Ƃ̔z��p�^�[���́AF. flexuosa �����ނ���Dipsastraea favus �i�~�L�N���C�V�ɗގ�����B����Ė{���F. flexuosa �́A�ʎ�ł���Ǝv���A���̕��ފw�I�ʒu�ɂ��Ă��X�Ȃ錟�����K�v�ł��낤�B�I�I�J���m�R�L�N���C�V�̘a����S���͖̂{��B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Favites yamanarii Yabe and Sugiyama, 1936
���}�i���J���m�R�L�N���C�V�i�V�́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Favites Link, 1807 �J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[5m)

|

䰂̒��a��1cm�O��B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B�����ł��A���ꂢ�ȑ��p�`�̌́A����䰕ǁA�K���������z��u�ǂ�F���ł���B
���i�̓����F�̂͑��p�`�ŁA���̔z��͊��S�ȃZ���I�C�h�^�B䰂̒��a��7�`1.4mm�����A1cm�O��̌̂������B䰕ǂ͌���1mm�O��ŁA�u�ǂɔ�ׂ�ƌ����k���Ɍ�����B�u�ǂ͒ʏ�S���܂Ŕ��B���A�قړ��������B䰕Ǐ㉏�œˏo�����A䰐S�Ɍ������Ċɂ₩�ɌX����B�u�Ǐ㉏�ɂ͑傫���ƊԊu�̑������������قړ��Ԋu�ł悭���B����B���̂���䰓��͐�����B�����Ɗu�ǂ̑��ʂɂ͏�����������̑������悭���B����B1���E2���u�ǂ́A�����܂ŒB����B3���u�ǂ������ɒB���邱�Ƃ͋H�����A���̂����߂��܂ŐL������B4���u�ǂ͒Z���A�L�тĂ���u�ǂ�2/3�قǁB�����͖��ĂŁA���a��1.5mm�قǂ̉~�`�`���ȉ~�`�B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ��A�J���I�Șp�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͖k�哌���B�{���Veron et al. (1977) ��Favites chinensis �V�i�L�N���C�V�̃V�m�j���ɂ���Ă���B�������{��̓V�i�L�N���C�V���䰌a���傫���A�V�i�L�N���C�V�i�{�K�C�h���f�ڎ�j�̂悤�Ƀv���R�C�h�^�ɂȂ邱�Ƃ�A�u�Ǔ����Ƀp����p����t���`������邱�Ƃ͂Ȃ��B�{���Favites abdita �J���m�R�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ƃ���������Ă���\��������A�����ł̐����ɂ��Ă͍Č������K�v�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Goniastrea favulus (Dana, 1846)
�q���E�l�J���m�R�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Goniastrea Milne Edwards and Haime, 1848 �R�J���m�R�L�N���C�V�� |
|

�B�e�F������^(���V�\�s��Óc�A���[12m)

|

䰂̒��a��3�`6mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂ��s�K���ɐ���オ������A�Q�̎������Œ���Ղ���傫���V�������肷�邱�Ƃ͏��Ȃ��B
��̕��̐F�ʂƓ����F���F�`�ΐF�܂��̓s���N�F�ŁA�W���F�ʂ̂��̂������B
���i�̓����F�̂͑��p�`�ŁA�����̔z��̓Z���I�C�h�^�`�����A���h���C�h�^�B����Q�̏�ł̌̂̑傫����`�͕s�����B䰂̒��a�́A��������̂��̂�3�`6mm�A������2�`3�����̂�9mm�`1.2cm���x�ɂȂ�B�u�ǂ�4���܂Ŕ��B���A䰂̒��a��6mm�قǂ̌̂�40���O��B�u�ǂ̏㉏�Ƒ��ʂɂ͐�[�̐���������Ɣ����ˋN�����ꂼ��悭���B����B����ē���ł��u�Ǖ\�ʂ��U�����Č����₷���B1�`3���u�ǂ͂قړ��������ŁA䰕Ǐ㉏����������ɒ���o�������ƁA䰒�Ɍ������Ăقڐ����ɗ������ށB4���u�ǂ͔����Z���A䰕Ǔ����ɂ킸���ɔ��B������x�B���̂���䰓��͐[���A���߂�1�`3���u�ǂƒZ��4���u�ǂ��K�����������݂ɕ���ł���悤�Ɍ�����B1���E2���u�ǂ͎����܂ŐL�����A�����̂����O���ɂ̓p����t���`�������B�����͏������A�s���Ă܂��͌����邱�Ƃ�����B䰕ǂ͔������k���ŁA�����⍂���͌Q�̂��Ƃɂ悭�����B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�A�ʒr���ʌ̊�Տ�ȂǂŌ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Goniastrea edwardsi �q���J���m�R�L�N���C�V�i�{�K�C�h���f�ڎ�j��G. pectinata �R�J���m�R�L�N���C�V�ƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��

|
|
| |
Goniastrea pectinata (Ehrenberg, 1834)
�R�J���m�R�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Goniastrea Milne Edwards and Haime, 1848 �R�J���m�R�L�N���C�V�� |
|

�B�e�F�����@�O(���V�\�s��v�ۍ`�A���[4m)

|

䰂̒��a��3�`6mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂɃR�u���~����̓ˋN���s�K���Ɍ`�����ꂽ��A�Q�̎������Œ���Ղ���傫���V�������肷�邱�Ƃ�����B
��̕��̐F�ʂƓ����F���F�`�ΐF�܂��̓s���N�F�ŁA�W���F�ʂ̌Q�̂������B
���i�̓����F�̂͑��p�`�ŁA�����̔z��̓Z���I�C�h�^�`���A���h���C�h�^�B����Q�̏�ł̌̂̑傫����`�͕s�����B䰂̒��a�́A��������̂��̂�4mm�`1cm�A������3�`6�����̂�1.6�`3cm���x�ɂȂ�B�u�ǂ�4���܂Ŕ��B���A䰂̒��a��8mm�قǂ̌̂�50���قǁB�u�Ǐ㉏�ɂ́A��[�ɕ����̔����ˋN���������G���ȋ������悭���B����B�����ɑΉ����āA�u�Ǒ��ʂł͔����ˋN���悭���B����B1�`3���u�ǂ͂قړ��������ŁA䰕Ǐ㉏�ł͓����Ɍ������Ă��[�u��ɒ���o�������ƁA䰒�Ɍ������Ăقڐ����ɗ������ށB4���u�ǂ͔����Z���A䰕Ǔ����ɂ킸���ɔ��B������x�B����āA���߂�1�`3���u�ǂƒZ��4���u�ǂ��K�����������݂ɕ���ł���悤�Ɍ�����B1���E2���u�ǂ͎����܂ŐL�����A�u�Ǔ����ɂ͔�������p����t���悭���B����B���̂���䰂͎��ۂقǂ͐[�������Ȃ��B�����̎���������䰂ł́A�אڂ��鎲���Ԃ̊u�ǂ͔������B�����͏������A�s���Ă܂��͌����邱�Ƃ�����B䰕ǂ͌����k���ŁA���[�u��ɒ���o����1�`3���u�ǂɂ���āA䰕Ǐ㉏�͊ۂ݂�ттĂ������������B�������A����Q�̓��ł̌����⍂���͕s�����ɂȂ�₷���B�Q�̂̒��Ŋ�ՂɌŒ����Ă��Ȃ����������̗��ʂ́AMerulina�T�U�i�~�T���S����Scapophyllia�I�I�T�U�i�~�T���S���̂悤�ɌQ�̎����Ɍ������ĕ��ˏ�ɔg�łB
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�A�ʒr���ʌ̊�Տ�ȂǂŌ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͂���܂�Goniastrea favulus �q���E�l�J���m�R�L�N���C�V�ƍ�������Ă������߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
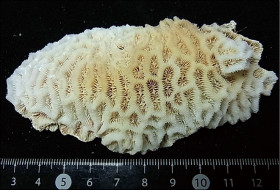
|
|
| |
Goniastrea retiformis (Lamarck, 1816)
�R�����L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Goniastrea Milne Edwards and Haime, 1848 �R�J���m�R�L�N���C�V�� |
|

�B�e�F�[���T�L(����q���剖���A���[2m)
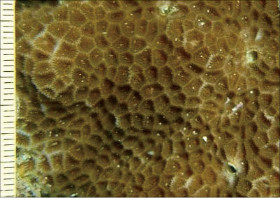
|

䰂̒��a��3�`6mm�B
|
|
�����`�F�핢��`�����Q�́B�傫�ȌQ�̂ł͕\�ʂ��s�K���ɐ���オ��B�Q�̎������Œ���Ղ���傫���V�����邱�Ƃ͏��Ȃ��B
��̕��̐F�ʂƓ����F���F�`�ΐF�܂��̓s���N�F�ŁA�W���F�ʂ̌Q�̂������B䰕Ǐ㉏����������Ɍ�����B
���i�̓����F�̂͑��p�`�ŁA䰂̒��a��3�`6mm�قǁA�Z���I�C�h�^�ɔz��B����Q�̏�ł̌̂̑傫����`�͕s���������A�̂����������߂ɂ��̈Ⴂ�͖ڗ����Ȃ��B�u�ǂ�4���܂Ŕ��B���A䰌a��4mm�قǂ̌̂ł�40���O��B�u�ǂ̏㉏�ɂ́A��[�ɕ����̔����ˋN���������������悭���B����B�����ˋN�͊u�ǂ̑��ʂɂ��悭���B����B1�`3���u�ǂ͂قړ��������ŁA䰕Ǐ㉏���玲���Ɍ������ċ}�X�ŗ������݁A1���E2���u�ǂ͎����܂ŒB����B1���E2���u�ǂ̓����ɂ́A�������͂ނ悤��6�`10�̂悭��������p����t�܂��̓p�����`�������B3���u�ǂ͎����߂��܂ŐL�т���̂����邪�A������1���E2���u�ǂ̔��������̒����B4���u�ǂ͔��ɔ����Z���A䰕Ǔ����ɂ킸���ɔ��B������x�B䰕ǂ��k���ŁA�����͌̂�Q�̂ɂ���ĕς��₷�����A���̏㉏�͏�ɔ�������Ă���B
�������F�g������̋����ʉ��A�ʎΖʂ��ʎΖʂ̐��[5m�Ȑ��A�ʌ��̊�Տ�ȂǂŌ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�FGoniastrea minuta �i�{�K�C�h���f�ڎ�j�ƍ�������Ă���\���������A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Hydnophora bonsai Veron, 1990
�{���T�C�C�{�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Hydnophora Fischer von Waldheim, 1807 �C�{�T���S�� |
|

�B�e�F���{�@��(���V�\�s�Y�c�p�A���[5m)
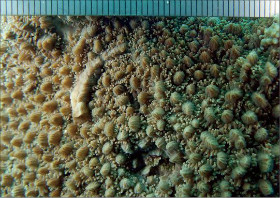
|

�����e�B�N����̒Z�a��2�`3mm�B
|
|
�����`�F�핢��`���Q�́B�Q�̕\�ʂɂ͒��a1.5�`2cm�A�������ő�5cm�قǂ̃R�u��`�~����̓ˏo�����s�K���ɔ��B���邱�Ƃ������B�܂��A�~���̐�[�͂���ɕ����ɕ��₷���B
��̕��̐F�ʂƓ����F�W���F�`�W�ΐF�B�����e�B�N���̕\�ʂƂ���ȊO�̕����̐F�ʂ͈قȂ邱�Ƃ������B���Ԃł��G���L�����Ƃ����邪�A�����e�B�N�����B��Č����Ȃ��قǂ܂ŐL�����邱�Ƃ͂Ȃ��B
���i�̓����F�Q�̕\�ʂ̓����e�B�N���ň�l�ɕ����A�̂̓n�C�h�m�t�H���C�h�^�ɔz��B�����e�B�N���͉~�`�`�ȉ~�`�ŁA��ł̒Z�a�͂��悻2�`3mm�A������3�`5mm�قǂɂȂ�B�����́A�ׂ����ɍ���������̂ŁA�Q�̕\�ʂ���悭�ˏo���Č�����B�Q�̎�����~���̐�[�������A�����̃����e�B�N�����A�Ȃ��ăR�������`�����邱�Ƃ͏��Ȃ��B�u�ǂ�3���܂łŁA�����e�B�N���̑��ʂɂ悭���B����B1���E2���u�ǂ́A�����e�B�N���̒������玲�����Ȃ��悤�ɔ��B����B3���u�ǂ͔��B�������A�ˋN�̑��ʂł킸���Ɍ�������x�B�u�Ǐ㉏�ɂ͋������A���ʂɂ�������ˋN�����ꂼ�ꔭ�B���邪�A�ǂ���������œ���ł͌����ɂ����B�����͊u�Ǔ�������L�т��g���x�L�����Ō`������邪�A����ł̊u�Ǔ����Ƃ̋�ʂ͍���ł���B
�������F�ʒr���ʌ�A�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�É������ɓ��E��ܓ����ʓ��ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Hydnophora exesa �g�Q�C�{�T���S��H. microconos �����E�L���E�C�{�T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���B����č����ł̖{��̐����ɂ��čČ�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Hydnophora exesa (Pallas, 1766)
�g�Q�C�{�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Hydnophora Fischer von Waldheim, 1807 �C�{�T���S�� |
|

�B�e�F���{�@��(���V�\�s�Y�c�p�A���[7m)

|

�����e�B�N����̒Z�a��3�`4mm�B
|
|
�����`�F�핢��`���Q�́B�Q�̕\�ʂɂ͒��a2.5�`5cm�A�������ő�10cm�قǂ̃R�u��`�~����̓ˏo�����s�K���ɔ��B���邱�Ƃ�����B�G��́A�����e�B�N�����F���ł��Ȃ��قǒ��Ԃł������L�т�B
��̕��̐F�ʂƓ����F�S�̓I�ɊD�F�����������F�`�ΐF�ŁA�Ԃ݂�тт����̂�������B
���i�̓����F>�Q�̕\�ʂ̓����e�B�N���ŕ����A�̔z��̓n�C�h�m�t�H���C�h�^�B�����e�B�N���͎�ɑȉ~���`�ŁA��ł̒Z�a��3�`4mm�A�����͍ő�5mm�قǂɂȂ�B�܂��A�����̃����e�B�N�����A�Ȃ��ăR�������`�����A�����̊Ԃ̌̂����A���h���C�h�^�ɔz�邱�Ƃ������B�����e�B�N���̑��ʂɂ�1�`3���u�ǂ����B����B1���E2���u�ǂ͒����A�����e�B�N���̒������玲���܂Ŕ��B���A�����ɋ߂��ق��ł͂������Ȃ�B3���u�ǂ͔����Z���A�����ɂ͒B���Ȃ��B�u�Ǐ㉏�ɂ́A��[�̐�����������悭���B����B�܂��u�ǂ̑��ʂɂ�������ˋN�����B���邪�������Ėڗ����Ȃ��B�����́A1���E2���u�ǂ̓�������L�т������̃g���x�L�����Ō`������邪�A����炪�A�����Ĕ�Ɍ�����ꍇ��A�����ꍇ������B
�������F�ʎΖʂł͐��[10m�Ȑ[�A��ʎΖʂł͐��[15m�Ȑ�Ō�����B�Օ��I�ȓ��p�ł�������B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Hydnophora exesa �g�Q�C�{�T���S�ƍ�������Ă��邽�߁A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Leptoria phrygia (Ellis and Solander, 1786)
�i�K���T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Leptoria Milne Edwards and Haime, 1848 �i�K���T���S�� |
|

�B�e�F������^(����q���n���̊≮�A���[2m)

|

�R���������Ԃ̋�����3�`4mm�B
|
|
�����`�F�핢��`�����Q�́B�傫�ȌQ�̂ł́A�\�ʂɑ傫�ȉ��݂�A���a�ƍ�������cm�ɂ���ԃR�u��`�~����̓ˏo�����s�K���Ɍ`������₷���B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA���F��ΐF�̌Q�̂�������B
���i�̓����F�̔z��͊��S�ȃ��A���h���C�h�^�B�����I�ɃZ���I�C�h�^�⏀���A���h���C�h�^�ɂȂ邱�Ƃ͂قƂ�ǂȂ��B䰕ǂɑ�������R�����͌���2�`3mm�A����2�`2.5mm�ŁA����R�����̒����Ԃ̋�����3�`4mm�B���������Ԍ��Ղ͕�2mm�����Ƌ������߁A����ł̓R����������������B�u�ǂ̓R�����ɒ�������悤�ɔ��B���A�قړ��Ԋu�Ŕz��B�u�ǂ̎����͕s���Ă����A��u�ǂ������A�����܂ŒB����̂ɑ��A�����u�ǂ͒Z���A�����܂ŒB���邱�Ƃ͂Ȃ��B�u�ǂ̓R���������œˏo���邪�A�㉏���s����邱�Ƃ͂Ȃ��A�����Ɋւ�炸�������悭�����B�u�ǂ̏㉏�ɂ͔����ȋ��������B����B�����̑��ʂƂ��̉������ɂ�����u�ǂ̑��ʂ�������ˋN�����B���邱�Ƃ�����B���ĂȎ����͌`������Ȃ����A�אڂ��鎲���Ԃɂ́A�k���Ȕ�܂��͖_��ˋN���`������A����炪������܂��͓_����ɔz��B
�������F�g������̋����J���I�ȏʎΖʂ̐��[10m�Ȑ�Ō�����B�܂����ʂ��̂悢�ʒr���ʌł�������B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Merulina ampliata (Ellis and Solander, 1786)
�T�U�i�~�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Merulina Ehrenberg, 1834 �T�U�i�~�T���S�� |
|

�B�e�F���{�@��(���V�\�s�Y�c�p�A���[3m)
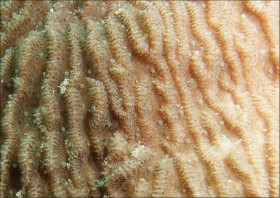
|

䰂̒��a��3�`6mm�B
|
|
�����`�F�Œ���Ղɉ����Đ�������핢��܂��͔���Q�́B�Q�̂̒��S�t�߂ł́A���a�ƍ������Ƃ���2�`3cm�قǂ̃R�u��̓ˏo�����s�K���Ɍ`������邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W�����F�A���邢�ΐF�܂��͔����s���N�F�B
���i�̓����F�̂�䰂̒��a��3�`6mm�قǂ̑��p�`�ŁA�̔z��̓��A���h���C�h�^�����A�Q�̂̒��S�t�߂�R�u��ˏo����ł́A�Z���I�C�h�^�ɂȂ邱�Ƃ�����B䰕ǂ͌���1�`1.5mm�قǁB�u�ǂ�3���܂Ŕ��B���邪�A3���u�ǂ������B�̌̂������B1���E2���u�ǂ�䰕Ǐ㉏�ł悭�ˏo����B�����̊u�ǂ͎����ɒB���邪�A�p����t�̔��B�͈����B3���u�ǂ�1���E2���u�ǂ��������Ĕ��ɒZ���A�����ɂ͒B���Ȃ��B�u�Ǐ㉏�ɂ́A��[�������˂��ꂽ�������A���ʂɂ͐�[������������ˋN�����ꂼ��悭���B����B���̂��ߓ���ł́A�u�ǂ������U�����Č�����B�����͏������A�u�Ǔ����Ƃ̋�ʂ��t���ɂ����B���A���h���C�h�^�ɘA�Ȃ��������̌̂́A1�̎��������L���邱�Ƃ͂Ȃ��A�אڂ��鎲���͕K����������u�ǂŊu�Ă��Ă���B�Q�̗��ʂ�3�`6mm�قǂ̊Ԋu�ŕ��ˏ�ɑ傫���g�ł��A���S�~��̃G�s�e�J�͌`������Ȃ��B�����œʏ�ɐ���オ���������ɂ́A6�`10�{���x�̖��ĂȘ]�������A���̏㉏�ɂ͌`��傫���̕s�����ȋ������悭���B����B����A�������ɂ́A�s�K���ɏ����ȍE���J�����Ƃ������B
�������F�ʒr�E��ʌ�A�g������̎ア�ʎΖʂ��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Scapophyllia cylindrica �I�I�T�U�i�~�T���S�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č�������K�v������B
* ��q�����L�^��

|
|
| |
Mycedium elephantotus (Pallas, 1766)
�E�X�J�~�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Mycedium Milne Edwards and Haime, 1851 �E�X�J�~�T���S�� |
|

�B�e�F��������(���V�\�s��Óc�A���[10m)
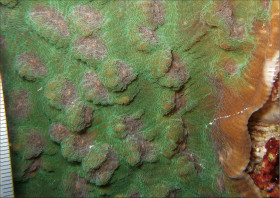
|

�̂͑����������B
|
|
�����`�F�핢��Q�́B�Ζʏ�ł͌Q�̎������t��ɒ���o�����Ƃ������B��ՂɌŒ����Ă���̂͌Q�̂̒��S�݂̂ŁA���̕����͊�Ղ���V�����Ă���B�����ł��A�̂��ˏo���A�Q�̎����������Ă���l�q���悭������B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Ԃ݂�тт����̂�D�F�����������́A���Ղ������Ȃ������̂������B
���i�̓����F�̂͂��ȉ~�`�`�ȉ~�`�A���a��5mm�`1cm�قǂ����A�o��r���܂��͒���̌̂͂�������傫���A�܂��͏������Ȃ�B�c�Q�̂ł́A�Q�̒��S�ɒ��a��1.5cm�قǂ̒��S�̂������邱�Ƃ�����B�̂̑����́A�Q�̎����������A��{�I�ɂ͒��S�̂��͂ނ悤�ɕ��ˏ�܂��͓��S�~��ɔz��B�������A�Q�̎����̗t�������A�K���I�Ȕz���F�����ɂ����B�̔z��̓v���R�C�h�^�����A�̂̉����ł�䰕ǂ������Ɠ������Ă���A���������z��ɂ͌����ɂ����B�Q�̎����ɂ����āA�����Ȍ̂ł�2���u�ǁE�]�܂ŁA�傫�Ȃ��̂ł�3���u�ǁE�]�܂Ŕ��B����B䰕ǂƋ����͔����A������0.5mm�����B1���E2���u�ǂ͌��߂ł悭�ˏo���A䰐S�̎����܂ŒB����B3���u�ǂ͔����A�����ɂ͒B���Ȃ��B�����������̂�1���E2���]�́A�����̂̓����܂��͗אڂ���ʌ̂�1���E2���u�ǁE�]�Ƃ��ꂼ��A������B3���]�́A�אڌ̂�3���u�ǁE�]�ƘA�����邱�Ƃ����邪�A�ʏ�͒f���I�B�u�ǁE�]�̏㉏�ɂ́A��[������������ȋ��������B����B�����ɂ́A�P���[�̂��̂�ו���[�̂��̂�������B�����͑ȉ~�`�ŁA���a��3mm�߂��܂łȂ邪�A�Q�̎����ɋ߂��̂قǔ��B�������s���āB�Q�̎����̕\�ʂƗ��ʂł́A1���E2���]��3���]���K�����������Ԋu�ŕ��ˏ�ɕ��ԁB�]�Ƙ]�̊Ԃ̋����ɍE�͊J���Ȃ��B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ̐��[20m�Ȑ�Ō�����B�J���I�ȓ��p�ł͂��ł������邱�Ƃ�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B��q���ł͂��܂Ɍ�����B
�����EVeron (1995)�ł̏o���L�^�F���iGoniastrea australensis�Ƃ��ċL�^�j

�������ʐ^�̌Q�̂Ƃ͕ʌQ��(�F�{���V�����[�A���[7m�ō̏W)�B
|
|
| |
Oulophyllia crispa (Lamarck, 1816)
�I�I�i�K���T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Oulophyllia Milne Edwards and Haime, 1848 �I�I�i�K���T���S�� |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[4m)

|

�R�����̍����͂悭�����B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�W���F�`�D�ΐF�B�R�����̒����͂�╽�R�Ɍ����A���̒����ɋ؏�̍a���`������₷���B
���i�̓����F�̔z��̓��A���h���C�h�^�ŁA�H�ɌQ�̒������Z���I�C�h�^�ɂȂ�B䰕ǂɑ�������R�����̊�ł̌�����1.5mm�`1.5cm�ƕω��ɕx�ނ��A�����͏��1mm�����Ŕ����B�R������������Q�̂ł́A�����߂��ŕs�K���Ɍ����J���B�R�����̍�����5mm�`1cm�ŁA�Q�̂��Ƃɂ悭�����B䰂̒Z�a�ɑ�������A����R�����̒����Ԃ̋�����1�`2cm�B�u�ǂ�4���܂Ŕ��B���A���Ԋu�Ő��R�ƕ��ԁB�u�ǂ̒����͎������ƂɈقȂ邪�A������4���u�ǂ���┖�������ő��͂قړ����Ɍ�����B1���E2���u�ǂ͎����܂ŒB����B�����̓����ɂ̓p����t�������邱�Ƃ�������̂̕s���āB3���u�ǂ́A�����̎�O�܂Řp�Ȃ��Ȃ���L�т�1���E2���u�ǂɗZ������B4���u�ǂ͒Z���A�R�����̓����ʂɂ킸���ɔ��B����̂݁B�u�Ǐ㉏�ɂ͑傫���ƍ����̂悭�����������ȋ��������R�ƕ��ԁB�]�́A�Q�̗̂��ʎ����Ɍ����A�u�ǂƓ��l�A�㉏�ɏ����ȋ��������B����B�����͉~�`�`���ȉ~�`�Œ��a2�`3mm�A1���E2���u�ǂ���L�т������̃g���x�L��������Ȃ�B�������Q�̂ɂ���ẮA�������s���Ă܂��͖����B�̌̂�������B�אڂ��鎲���Ԃɂ́A��������u�ǂ܂��͑���̃g���x�L�������`�������B
�������F>�J���I�ȏʎΖʂ̐��[10m�Ȑ[�A�܂��͑傫�Șp���̐��[5m�Ȑ[�Ō�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�����EVeron (1995)�ł̏o���L�^�F���iGoniastrea deformis�Ƃ��ċL�^�j

|
|
| |
Oulophyllia sp.
�g�K���I�I�i�K���T���S�i�V�́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Oulophyllia Milne Edwards and Haime, 1848 �I�I�i�K���T���S�� |
|

�B�e�F�����@�O(���V�\�s�Y�c�p�A���[5m)

|

�R�����̍����͕s�����B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�D���F�`�D�ΐF�ŁA��̕��͓����Ɍ�����B�R���������́A��u�Ǒ��������s�K���ɓˏo���Č�����B
���i�̓����F�̔z��̓Z���I�C�h�`���A���h���C�h�^�B�R�����̊�ł̌�����3mm�`1cm�ŕs�����A��̂ق��������A�����Ɍ������Ĕ����Ȃ�X��������B�������s�����ŁA�Ⴂ�Ƃ���ł�5mm�قǁA�悭�ˏo�����Ƃ���ł�1cm����B����R�����̒����Ԃ̋����́A�����Ƃ����5mm�A�L���Ƃ���ł�3cm�߂��ɒB����B�����ˏo�����R�����̒����߂��ł́A�E���J������傫�ȗڂ��ł����肷��B�u�ǁE�]�͒ʏ�4���܂Ŕ��B���A��̂��̂قǒ��������A�R���������ō����ˏo����B�̂ɂ���ẮA1���u�ǂ�䰂̏����䰓��Ɍ������č�������o���A���̑��ʂ�3���E4���u�ǂ����B���邱�Ƃ�����B1���E2���u�ǂ͎����܂ŒB����B�u�Ǔ����ł̃p����t�̔��B�͈����B3���E4���u�ǂ͎����ɒB�����A��������O�Řp�Ȃ��Ȃ����u�ǂɗZ������B�u�Ǐ㉏�ɂ͑傫���̑��������������R�ƕ��сA���̑��ʂƉ������̊u�Ǒ��ʂɂ͏����Ȟ���̓ˋN���`�������B�]�́A�Q�̗̂��ʎ����ɂ悭���B���邪�A�u�ǂƈ���Ď����ɂ��Ⴂ���s���ĂŁA�����̔��B�������B�u�ǂƓ��l�A���̏㉏�ɂ͏����ȋ��������B����B�����͒��a��2�`4mm�̉~�`�`�ȉ~�`�ŁA1���E2���u�ǂ̓�������L�т������̃g���x�L�����ɂ���Č`�������B�̔z�����A���h���C�h�`���A���h���C�h�^�̂Ƃ���ł́A�אڂ��鎲���Ԃ�1���u�ǂƓ����x�܂��͂���ȏ�ɔ�������u�ǂ������邱�Ƃ������B
�������F�J���I�ȏʎΖʂ̐��[10m�Ȑ[�܂��͑傫�Șp���̐��[5m�Ȑ[�Ō�����
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�V�̘a���́A��u�ǂ��悭�ˏo���A�R�����̒������e�������邱�ƂɗR������B
* ��q�����L�^��

|
|
| |
Paragoniastrea australensis (Milne Edwards and Haime, 1857)
�E�l�J���m�R�L�N���C�V�i�V�́j |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Paragoniastrea Huang, Benzoni and Budd, 2014 �E�l�J���m�R�L�N���C�V���i�V�́j |
|

�B�e�F������^(����q���n���̊≮�A���[10m)

|

䰂̒��a��1�`1.5cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�Ê��F�`�W���F�A�܂��͗ΐF�B�D�F���������Q�̂���Ղ������Ȃ����Q�̂������B
���i�̓����F�̂͑��p�`�ŁA2�`3�̌̂��A�Ȃ��ăZ���I�C�h�^�`�����A���h���C�h�^�ɔz��B䰂̒��a��1�`1.5cm�A�����̌̂��A�Ȃ����Ƃ���ł̒��������̒����͍Œ���3cm�قǂɂȂ�B䰕ǂ́A������������͔����B�������A�����͏��0.5mm�����łǂ̌Q�̂ł��قړ��������A��͌�����1�`5mm�ŕω����A�����Q�̓��ł��ꏊ�ɂ���Č������قȂ邱�Ƃ�����B䰕ǂ̍�����5�`8mm�قǁB���̂���䰓��͐[�����A䰕ǂ̌`��ɂ���Ď��ۂقǂ͐[�������Ȃ��B�u�ǂ�4���܂Ŕ��B���A���̌����͎����ɂ�炸�قړ����ŁA䰌a��1.2cm�قǂ̌̂ł́A�u�ǐ���36�`40�����x�B������䰕Ǐ㉏�ō����ˏo���邱�Ƃ͂Ȃ��A䰕Ǐ㉏����͓����ʂɉ����ċ}�X�ŗ������ށB1���E2���u�ǂ͎����܂ŒB���A�����ɂ̓p����t�܂��̓p�����悭���B����B3���u�ǂ̒����͌̂ɂ���ĈقȂ邪�A�L�тĂ���̊u�ǂ̃p����t�܂��̓p���̎�O�܂ŁB4���u�ǂ́A䰕Ǔ�������킸���ɒ���o���̂݁B�u�Ǐ㉏�ɂ́A��[����������������R�Ɣz��B�������ʂ��炻�̉����̊u�Ǒ��ʂɂ́A��[���悭����������ˋN���悭���B����B������2�`3mm�̉~�`�܂��͑ȉ~�`�Ŗ��āB
�������F�ʎΖʂ̐��[20m�Ȑ[���ʎΖʂ̐��[15m�Ȑ�Ō�����B�Օ��I�ȓ��p�ł́A���[5m�Ȑ�ł����邱�Ƃ��ł���B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Paragoniastrea sp. �ƍ�������Ă���\�������邽�߁A�{��̍����ł̐����ɂ��Ă͍Č������K�v�B
* ��q�����L�^��
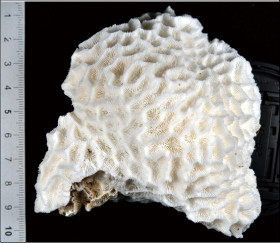
|
|
| |
Paragoniastrea deformis (Veron, 1990)
�~�_���J���m�R�L�N���C�V |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Paragoniastrea Huang, Benzoni and Budd, 2014 �E�l�J���m�R�L�N���C�V���i�V�́j |
|

�B�e�F�����@�O(���V�\�s��Óc�A���[10m)

|

䰂̒��a��5mm�`1cm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F��ɗΐF�A�Ê��F��Ԋ��F�B䰕Ǒ��ʂ������ۂ��A���Ղ������ۂ��Ȃ����̂͐����ł��悭�ڗ��B
���i�̓����F�̂͂��p�����~�`�`���p�`�A䰂̒��a��5mm�`1cm�ŁA7�`8mm�̂��̂������B�̂̓v���R�C�h�^�܂��̓Z���I�C�h�^�ɔz�A�̊Ԃɂ́A�Ǐ�܂��͍a��̑��їނ̐��ǂ��s�K���ɔ��B����B�u�ǁE�]��4���܂Ŕ��B����B������4���u�ǂ́A䰕Ǐ㉏���炻�̓����ɂ킸���ɔ��B���邩�����B�̏ꍇ���قƂ�ǁB�u�ǐ���䰌a8mm�قǂ̌̂�30�`40���ɂȂ�B�]�́A�̊Ԃ��悭���ꂽ�����Ō`������邱�Ƃ����邪�A�Z���Ėڗ����Ȃ��B�u�ǂƘ]�̏㉏�ɂ͋������A�����Ɗu�ǂ̑��ʂɂ͔����ˋN���悭���B����B1���E2���u�ǂ͊u�Ǐ㉏�Ŕ�����₷���A������������䰓��ɒ���o�������ƁA�}�X�ŗ������݂Ȃ��玲���܂ŒB����B�����̊u�ǂ̓����ɂ̓p����t�����B���邱�Ƃ������B�����͖��ĂŁA1���E2���u�ǂ���L�т��g���x�L�����Ō`�������B3���u�ǂ͒������s�����ŁA�����߂��܂ŐL�т���̂�����A�u�Ǔ����ɂ킸���ɔ��B������̂�������B䰕ǂ͌��߂ŁA�킸���ɏ���ɓˏo����B�����͒��a��1.2mm�O��̉~�`�܂��͑ȉ~�`�ŁA���ĂȌ̂�����Εs���ĂȂ��̂�����B
�������F�ʎΖʂł͐��[20m�Ȑ[�ŁA��ʎΖʂł͐��[15m�Ȑ�Ō�����B
�����ł̕��z�F��t���َR�E���茧��ܓ����ʓ��ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{��̃^�C�v�Y�n�͘a�̎R�����{�B
*** ����܂Ŏ�q���Ŋm�F����Ă��Ȃ���
|
|
| |
Paragoniastrea sp.
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Paragoniastrea Huang, Benzoni and Budd, 2014 �E�l�J���m�R�L�N���C�V���i�V�́j |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[8m)

|

䰂̒��a�͒ʏ�7�`8mm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�Ê��F�`�Ԋ��F�B
���i�̓����F�̂͂��p�����~�`�`���p�`�A䰂̒��a��5mm�`1cm�ŁA7�`8mm�̂��̂������B�̂̓v���R�C�h�^�܂��̓Z���I�C�h�^�ɔz�A�̊Ԃɂ́A�Ǐ�܂��͍a��̑��їނ̐��ǂ��s�K���ɔ��B����B�u�ǁE�]��4���܂Ŕ��B����B������4���u�ǂ́A䰕Ǐ㉏���炻�̓����ɂ킸���ɔ��B���邩�����B�̏ꍇ���قƂ�ǁB�u�ǐ���䰌a8mm�قǂ̌̂�30�`40���ɂȂ�B�]�́A�̊Ԃ��悭���ꂽ�����Ō`������邱�Ƃ����邪�A�Z���Ėڗ����Ȃ��B�u�ǂƘ]�̏㉏�ɂ͋������A�����Ɗu�ǂ̑��ʂɂ͔����ˋN���悭���B����B1���E2���u�ǂ͊u�Ǐ㉏�Ŕ�����₷���A������������䰓��ɒ���o�������ƁA�}�X�ŗ������݂Ȃ��玲���܂ŒB����B�����̊u�ǂ̓����ɂ̓p����t�����B���邱�Ƃ������B�����͖��ĂŁA1���E2���u�ǂ���L�т��g���x�L�����Ō`�������B3���u�ǂ͒������s�����ŁA�����߂��܂ŐL�т���̂�����A�u�Ǔ����ɂ킸���ɔ��B������̂�������B䰕ǂ͌��߂ŁA�킸���ɏ���ɓˏo����B�����͒��a��1.2mm�O��̉~�`�܂��͑ȉ~�`�ŁA���ĂȌ̂�����Εs���ĂȂ��̂�����B
�������F��ʎΖʂ̐��[15m�Ȑ�Ō�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Paragoniastrea australensis �E�l�J���m�R�L�N���C�V��Favites abdita �J���m�R�L�N���C�V�i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���A�����ł̐����̍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Pectinia lactuca (Pallas, 1766)
�X�W�E�~�o�� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Pectinia de Blainville, 1830 �X�W�E�~�o���� |
|

�B�e�F�����C��(���V�\�s�Y�c�p�A���[6m)

|

�R�����͔����A�����ˏo����B
|
|
�����`�F�핢��`�t��Q�́B�悭�����オ���Ē����A�Ȃ����R���������B���邽�߁A�Q�̂̊O�ς̓h�[���^�`�����`�Ɍ�����B
��̕��̐F�ʂƓ����F�ΐF�`���F�B�Ԃ݂�тт��Q�̂≩�F���ۂ��Q�̂�������B
���i�̓����F�Q�̎����̌̂́A���a��1�`3cm���x�B�c�Q�̂ł́A�傫�߂̒��S�̂�1����A�����̏����Ȏ����̂����͂��݂͂Ȃ���z�A�����I�ɂ͂���炪���S�~��ɔz�邱�Ƃ�����B�Q�̂̐����ɔ����A�V���Ɍ`�����ꂽ�����̂̓��A���h���C�h�^�ɔz��B���S�̂�1���E2���u�ǂ́A�������痣���ɂ�č����ˏo����B�����̊u�ǂ́A�Q�̂̐����ɔ����Ď����̂�䰕ǂ̈ꕔ�ƂȂ�A���ɂ悭�A�������R�������`������B�R�����̍����́A�Q�̒��S�ł�4cm���x���邪�A�Q�̎����Ɍ������ɂ�ĒႭ�Ȃ�B�����̂̊u�ǂ́A5���܂Ŕ��B����B���S�̂Ɠ��l�ɁA1���E2���u�ǂ̎����͍����ˏo����B�אڂ�������̂́A�悭�ˏo�����R����������Ō݂��̊u�ǁE�]�̈ʒu���悭��v����B�܂��A����炪��v�����ӏ���䰕ǂɂ́A�s�K���ɍE���J�����Ƃ������B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ̐��[20m�Ȑ�Ō�����B�ʒr���ʌ̐ł��H�Ɍ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋ɂ߂ċH�B
�⑫�F�{���Pectinia paeonia ���[�X�E�~�o���i�{�K�C�h���f�ڎ�j��Physophyllia ayleni �E�~�o���ƍ�������Ă���A�����ł̐������Č�������K�v������B
* ��q�����L�^��
|
|
| |
Physophyllia ayleni Wells, 1935
�E�~�o�� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Physophyllia Duncan, 1884 �E�~�o���� |
|

�B�e�F�����@�O(���茧���s�Y�A���[6m)

�������ʐ^�Ɠ����Q�̂Ŏ�q���Y�ł͂Ȃ��B |

䰂̒��a��9mm�`1.5cm�B
|
|
�����`�F��ɔ핢��Q�́B�R�������悭���B���ėt��Q�̂ɂȂ���̂�����B
��̕��̐F�ʂƓ����F�D�F�����������ΐF�`���F�B���Ղ����F��ԐF�Ȃǂ̑N�₩�ȐF�ʂɂ��Ȃ�B
���i�̓����F�Q�̎����͉̌̂~�`�`�ȉ~�`�Œ��a��9mm�`1.5cm�قǁB�����Ȕ핢��Q�̂ł́A�傫�Ȓ��S�̂�F���ł���B�����̌̂́A�����������Ȃ���A���S�̂��͂ނ悤�ɕ��ˏ�܂��͓��S�~��ɔz��B�������̂̑傫����z�u���s�����ŁA�Q�̕\�ʂ̋N�����傫�����߁A�K���I�ɔz��悤�Ɍ����ɂ����B�̔z��͊�{�I�Ƀv���R�C�h�^�����A�̂����Q�̎������������߁A�T�^�I�ȃv���R�C�h�^�ɂ͌����ɂ����B�����̂́A�����Ȃ��̂ł�2���u�ǁE�]�܂ŁA�傫�Ȃ��̂̈ꕔ��3���u�ǁE�]�����B���A�Q�̎����ł͂ǂ��ł�3���]�����B����B1�`3���u�ǁE�]�̌����͂قړ����ł悭�ˏo���邪�A�ˏo�̓x���͒�̂��̂قnj����B1���E2���u�ǂ�䰕Ǐ㉏�Ń��[�u��ɂȂ��Ă���A�����܂ŒB����B3���u�ǂ̓��[�u��ɂȂ炸�A�����ɂ͒B���Ȃ��B1���E2���]�́A���̌̂̓����܂��͗אڂ���ʂ�2�̂�1���E2���u�ǁE�]�Ƃ��ꂼ��A������B�����͑ȉ~�`�ŁA���a5mm���x�܂łȂ邪�A�Q�̎����ɋ߂��̂قǔ��B�������s���āB�אڂ���]�̊Ԃɂ́A�ΊD���̖A��g�D���s�K���Ɍ`�������B����䰂̂����O���ł́A����炪�悭���B���邱�Ƃ�䰕ǂ��`������Ă���l�q���悭������B�Q�̎����̕\�ʂƗ��ʂł́A1�`3���]���K�����������Ԋu�����ˏ�ɕ��ԁB���������̘]�Ƙ]�̊ԂɍE���J�����Ƃ͂Ȃ��B
�������F�Օ��I�ő������p���̐��[5�`15m�Ō�����B
�����ł̕��z�F���茧�Δn�`�F�{���V�����[�A�É������ɓ��`���m�������B��q���ł͋L�^����Ă��Ȃ��B
�⑫�F�{��̃^�C�v�Y�n�͒��茧�Δn�̉\��������B�{���Pectinia paeonia ���[�X�E�~�o���i�{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���A�����ł̐������Č�������K�v������B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Platygyra daedalea (Ellis and Solander, 1786)
�q���m�E�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Platygyra Ehrenberg, 1834 �m�E�T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[2m)

|

�R���������Ԃ̋�����5�`7mm�B
|
|
�����`�F���Q�́B�Q�̕\�ʂɓˏo�����`������邱�Ƃ͋H�B�ʒr�ł͒��a1m���鋐��Ȕ�����Q�̂��`������B
��̕��̐F�ʂƓ����F�W���F�`���F�܂��͗ΐF�B�אڌ̂̋��E�ɂ�����R���������ł́A��̕\�ʂɍׂ��؏������镔��������B
���i�̓����F�̔z��̓��A���h���C�h�^�ŁA�����I�ɃZ���I�C�h�^�⏀���A���h���C�h�^�ɂȂ�B�R�����̌�����4mm�قǂɂȂ邱�Ƃ����邪�ʏ��2m�����Ŕ����B�R�����͍���5mm�قǂŁA�������猩��Ƃ��̒����߂��ŕs�K���ɍE���J���B����R�����̒����Ԃ̋�����5�`7mm�B���������Ԍ��Ղ̕���3�`4mm�ƍL�����߁A����ł̓R�����������A���Ղ��L��������B�u�ǂ�3���܂Ŕ��B���A�R�����ɒ�������悤�ɋK�����������Ԋu�Ŕz��B1���E2���u�ǂ͒����A�R���������ł悭�ˏo���A�����܂ŒB����B3���u�ǂ͒Z���A���܂�ˏo�����A�����܂ŒB���Ȃ��B�u�Ǐ㉏�ɂ͑傫���̕s�����ȋ������悭���B����B�����́A1���E2���u�ǂ̓�������L�т������̃g���x�L�����Ō`�������B������A������悤�ɁA������܂��͑����_��̓ˋN���`������邱�Ƃ�����B
�������F�ʒr���ʌŌ�����B�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�ł������邱�Ƃ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F��q���Ȗk�ł́A�{���Platygyra contorta �~�_���m�E�T���S�ƍ�������Ă���\�������邽�߁A�{��̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Platygyra cf. daedalea (Ellis and Solander, 1786)
�a���Ȃ� |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Platygyra Ehrenberg, 1834 �m�E�T���S�� |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[4m)
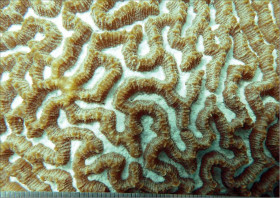
|

�R���������Ԃ̋�����5mm�O��B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F�W���F�`���F�܂��͗ΐF�B�������A���Ղ�1���E2���u�ǂ̏㉏�͏�ɔ����ۂ��Ȃ�B
���i�̓����F�̔z��̓��A���h���C�h�^�ŁA�����I�ɃZ���I�C�h�^�⏀���A���h���C�h�^�ɂȂ�B�R�����̌����͊���璸���܂ŋψ�ŁA1mm�قǂ����Ȃ��B�R�����̍�����5mm�قǂŁA�������猩��Ƃ��̒����߂��ɕs�K���ɍE���J���B����R�����̒����Ԃ̋�����5mm�O��ŁA���������Ԍ��Ղ̕���4mm�O��B�u�ǂ�3���܂Ŕ��B���A1���E2���u�ǂ͎����܂ŒB����B�܂������͔��B���ɂ������A1���E2���u�ǂ̓������킸���ɔ��������A�p����t�����B�����肷��X���ɂ��邽�߁A���̈ʒu�͔F�����₷���B�u�Ǐ㉏�ɂ͑傫���Ⓑ���̈قȂ鋘�������B����B����1���E2���u�ǂ̋����́A�R�����̒����߂��ŏ���ɂ悭�L������B
�������F�ʎΖʂ��ʎΖʂ̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Platygyra daedalea �q���m�E�T���S�AP. sinensis �V�i�m�E�T���S��Leptoria irregularis �~�_���i�K���T���S�i�����3��͑S�Ė{�K�C�h���f�ڎ�j�Ȃǂƍ�������Ă���\��������B�����ł̐����ɂ��Ă͍X�Ȃ錟�����K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Platygyra contorta Veron, 1990
�~�_���m�E�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Platygyra Ehrenberg, 1834 �m�E�T���S�� |
|

�B�e�F�i�c�r��(���V�\�s��v�ۍ`�A���[8m)

|

�R���������Ԃ̋�����4�`8mm�B
|
|
�����`�F�핢��`�����Q�́B�Q�̎������߂���オ������A�Q�̕\�ʂɒ��a�ƍ�����3�`4cm�قǂ̃R�u��ˏo�����s�K���Ɍ`�����ꂽ�肷�邱�Ƃ�����B
��̕��̐F�ʂƓ����F�W���F�`���F�܂��͗ΐF�B�R�����Ɗu�ǂ̏㉏�����������邢�F�ɂȂ�₷���B�g����ł́A��̂��R���������߂��̊u�Ǐ㉏�ŏ�����ɖc��݁A�����Ɍ����邱�Ƃ�����B
���i�̓����F�̔z��̓Z���I�C�h�`�����A���h���C�h�^�ŁA�H�ɌQ�̂̈ꕔ�����A���h���C�h�^�ɂȂ邱�Ƃ�����B�R�����͌���1�`3mm�A����3�`7mm�A����R�����̒����Ԃ̋�����4�`8mm�ƁA����Q�̏�ł��s�ψ�ɂȂ�₷���B�������Q�̓ˏo���̌̂͊T���đ傫���A����Ɍ������ă��b�p��ɊJ�����悤�Ɍ�����B�R�������悭�ˏo�����Ƃ���ł́A���̒����߂��ōE���J������A�傫�ȗڂ��ł����肷�邱�Ƃ�����B3���u�ǂ܂Ŕ��B���A�㉏�ɂ͑傫���̕s�����ȋ������悭���B����B1���E2���u�ǂ͎����ɒB���A�������͂ނ悤�Ƀp����t�����B���邱�Ƃ�����B�܂��A�R���������߂��ł́A�����������L�тāA���s�K���ȕ����������₷���B���̂��ߓ�������ɔ�ׂ�ƁA�Q�̕\�ʂ��k���(�����)������B3���u�ǂ͒������s�����ŁA������O�܂ł悭���B������̂�A�R�����̑��ʂɂ킸���ɔ��B������̂�����B�����͖��ĂŁA���a1�`1.5mm�̉~�`�A1���E2���u�ǂ̓�������L�т������̃g���x�L�����Ō`�������B
�������F���Օ��I�ȏʎΖʂ̐��[10�`20m�t�߁A���Օ��I�Ȋ�ʎΖʂ���p�̐��[10m�Ȑ�Ō�����B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͂��܂Ɍ�����B
�⑫�F�{���Paragoniastrea russelli ( ���w����Favites russelli �Ŗ{�K�C�h���f�ڎ�) �ƍ�������Ă��邪�A����͑S���̕ʎ�ł���B����č����ł̐����ɂ��čČ������K�v�ł���B
* ��q�����L�^��

|
|
| |
Scapophyllia cylindrica Milne Edwards and Haime, 1849
�I�I�T�U�i�~�T���S |
Meruliniidae�@Verrill, 1865 �T�U�i�~�T���S��
Scapophyllia Milne Edwards and Haime, 1848 �I�I�T�U�i�~�T���S�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[6m)
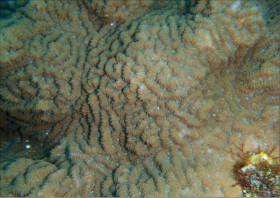
|

䰂̒��a��2�`5mm�B
|
|
�����`�F�핢��`�����Q�́B���a�ƍ�������cm�قǂ̉~����̓ˏo�����s�K���Ɍ`������邱�Ƃ������B�Q�̎����͌Œ���Ղ���킸���ɗV�����邪�A����ɂ߂��ꂽ��傫���g�ł����肷�邱�Ƃ͂Ȃ��B
��̕��̐F�ʂƓ����F�������F��ΐF�A�s���N�F�������B
���i�̓����F�̂́A䰂̒��a��2�`5mm�قǂ̑��p�`�B�̔z��͎�Ƀ��A���h���C�h�^�����A�Q�̂̒��S�߂���~�����ł͈ꕔ�Z���I�C�h�^�ɂȂ邱�Ƃ�����B�u�ǂ�3���܂Ŕ��B����B1���E2���u�ǂ͌������A䰕Ǐ㉏�ł͔����Ȃ�A�悭�ˏo����B�����̊u�ǂ͎����܂ŒB���A�����̂����O���ɂ̓p����t�����B���₷���B3���u�ǂ͔����A䰕Ǔ����ɉ����Ă킸���ɔ��B���邾���ŁA�����܂ŒB���邱�Ƃ͂Ȃ��B�u�ǂ̏㉏�ɂ́A��[�����A���˂��ꂽ�������A���ʂɂ͐�[������������ˋN�����ꂼ��悭���B����B���̂��߁A�u�ǂ͎��ۂ��������A�U�����Č�����B�����͏������A1���E2���u�ǂ̓�������L�т�2�`3�{�̃g���x�L�����Ō`������邪�A�����B�̌̂������B�܂��A���A���h���C�h�^�ɔz�������̌̂�1�̎��������L���邱�Ƃ͂Ȃ��A�אڂ��鎲���Ԃɂ͔�������u�ǂ����݂���B�R�����͌���1�`1.5mm�قǂŁA�r���ɍE���J�����Ƃ͂Ȃ��B�Q�̗��ʂ́A������2�`5mm�قǂ̊Ԋu�ŕ��ˏ�ɂ�邭�g�ł��Ă���A���̕\�ʂɓ��S�~��̃G�s�e�J�͌`������Ȃ��B�����œʏ�ɐ���オ���������ɂ́A�A�����̈����]��5�`8�{���x�����A���̏㉏�ɂ͌`��傫���̈قȂ鋘�����U�݂���B�܂��A����Ő���ɉ������ł́A�s�K���ɍE���J�����Ƃ�����B
�������F�ʎΖʂ̐��[5�`15m�Ō�����B�g�Q�̎ア�ʉ��`�ʎΖʏ㕔�ł������邱�Ƃ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Merulina ampliata �T�U�i�~�T���S�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č�������K�v������B
* ��q�����L�^��

|
|
| |
Pocillopora damicornis (Linnaeus, 1758)
�n�i���T�C�T���S |
Pocilloporidae Gray, 1840 �n�i���T�C�T���S��
Pocillopora Lamarck, 1816 �n�i���T�C�T���S�� |
|

�B�e�F�o�H���q(���V�\�s��v�ۍ`�A���[4m)

|

��3���}�͑�1�E��2���}�̐�[�ő����B
|
|
�����`�F�ő��R�����{�[�X��`�ő���Q�́B�T���S�ʂ̏ʒr���ʌł͂��ꂢ�ȃh�[���^�`�����`�ɂȂ�₷���B�������A�ʉ���ʎΖʂ̐Ȃǂ̔g������̋����Ƃ���ł́A�H�ɞ���Q�̂ɂȂ邱�Ƃ�����B�Q�̂̍�����10cm���邱�Ƃ͋H�B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA䰓��͔Z���F�ɂȂ�B
���i�̓����F��3���}�܂Ŕ��B�B��1���}�́A����f�ʂ��Z�a8mm�`1cm�̑ȉ~�`�ŁA�}�̒���4�`5cm�̂��̂������B��2���}���ȉ~�`�ŁA�Z�a6mm�O��A�}�̒���1.5�`2cm�B�C�{��̓ˋN���ɑ��������3���}�̊�ł́A���f�ʂ͉~�`�A���a2�`4mm�B��3���}�̒�����2�`5mm�̂��̂��������A6mm�ȏ�ɐL�т����̂�������B��3���}�́A��1�E��2���}�̐�[���ӂő��������A��3���}�Ԃ̋����͂ق�3mm�����B�̂͒��a1mm�O��̂��̂������B��1���}�̉�����Q�̊�͉̌̂~�`�ŁA�����̊Ԋu�͗����B����A��1���}�̏�����2�E��3���}�̌̂́A�p�����~�`�`���p�`�ŁA�אڌ̂�䰕ǂ̈ꕔ�����L����B�u�ǂ⎲���̔��B�͈������A�ꕔ�̌̂ł́A�ׂ��j��`�Z�����1���E2���u�ǂ�A�Z���~����`�Ⴂ�ȉ~�u��̎������F�߂��邱�Ƃ�����B�����\�ʂɂ́A�ׂ����G���ȒP���[����ו���[�������B����B
�������F�ʒr���ʌΓ��Ō�����B�g�Q�̎ア�ʎΖʂ��ʎΖʂł����[10m�Ȑ�Ō����邱�Ƃ�����B
�����ł̕��z�F���{�E�ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{��͂���܂�Pocillopora acuta �i�{�K�C�h���f�ڎ�j�ƍ�������Ă������߁A�����ł̗���̐������Č�������K�v������B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FS. pistillata�Ƃ��ċL�^�����\������
|
|
| |
Pocillopora eydouxi (Milne Edwards and Haime, 1860)
�w���W�J�n�i���T�C�T���S |
Pocilloporidae Gray, 1840 �n�i���T�C�T���S��
Pocillopora Lamarck, 1816 �n�i���T�C�T���S�� |
|

�B�e�F�쑺�b��(���V�\�s��Óc�A���[2m)

|

�אڂ����3���}�̊Ԋu�͍L��������B
|
|
�����`�F����`�R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�ŁA���F�A�s���N�F�⎇�F�Ȃǂ̔h��ȐF�ʂɂȂ�Q�̂�����B
���i�̓����F��3���}�܂Ŕ��B�B��1���}�̊T�ς́A���`�̑��A�ȉ~������`�ȂǕω��ɕx�ށB��1���}�́A����f�ʂ��Z�a1.8cm�O��̑ȉ~�`�ŁA�}�̒���10cm�ȏ�̂��̂��قƂ�ǁB��2���}�̊T�ς͞��`�܂��͑ȉ~���`�~����A����f�ʂ͒Z�a1�`1.8cm�B�}�̒����́A����炪�t�������1���}�����`��`�̏ꍇ��3cm�����ŒZ�����̂��������A�ȉ~����̏ꍇ��10cm�ȏ�ɂȂ�B��3���}�͑�1�E��2���}�̕\�ʂɖ��ՂȂ����B����B��3���}�͉~�`�ŁA��̒��a�Ǝ}�̒������Ƃ���2�`2.5mm�Ə����Ȃ��̂���A�ȉ~�`�ŒZ�a2.5�`3.5mm�A����3�`5mm�ɒB�����^�̂��̂܂ł���B��3���}�Ԃ̋�����2�`4mm�ŁA�����̒Z�a�ȏ�̊Ԋu���Ă���Ƃ�������Ȃ��Ȃ��B�̂͒��a1mm�O�ォ��菬�����~�`�ŁA�݂��ɗ���ĕ��z����B�������e���}�̐�[�ł́A�p�����~�`�`���p�`�ɂȂ��Ė��ڂ��A�אڌ̂�䰕ǂ̈ꕔ�����L����B1���E2���u�ǂƎ����͂悭���B����B�u�ǂ͑����j��A1���E2���u�ǂ̒����͂��ꂼ��1/2R��1/3R�قǁB�����͉~���`��M��ŁA����ł��F���ł���B�����\�ʂɂ͒P���[����ו���[�������B����B
�������F�T���S�ʈ�ł͕��ʂɌ����A���ɔg������̋����ʉ�����ʎΖʂ̐�(���[5m�Ȑ�)�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Pocillopora meandrina �`�������n�i���T�C�T���S�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
** ����̒����ł͊m�F�ł��Ȃ������������EVeron(1995)�ł͎�q���Ŋm�F����Ă����
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Pocillopora meandrina Dana, 1846
�`�������n�i���T�C�T���S |
Pocilloporidae Gray, 1840 �n�i���T�C�T���S��
Pocillopora Lamarck, 1816 �n�i���T�C�T���S�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[8m)

|

�אڂ����3���}�̊Ԋu�͋���������B
|
|
�����`�F����`�R�����{�[�X��Q�́B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�ŁA���F�A�s���N�F�⎇�F�Ȃǂ̔h��ȐF�ʂɂȂ�Q�̂�����B
���i�̓����F��3���}�܂Ŕ��B�B�ʏ�A��1�E��2���}�̊T�ς͞��`�ŁA����f�ʂ͑ȉ~�`�̂��̂������B������猩��ƁA��2���}���ʂ̈ꕔ����1���}�ɗZ�����A�����̏㉏�������֍s���Č����邱�Ƃ�����B��1�E��2���}�̊�ł̒Z�a�͂��ꂼ��1.5cm�O���1.3�`1.5cm�قǂŁA�}�̒����͂Ƃ���3cm�ȏ�ɐL�т�B��3���}�͑�1�E��2���}�̕\�ʂɖ��ՂȂ����B���A����f�ʂ͉~�`�`�ȉ~�`�A���a�܂��͒Z�a2�`3.5mm�A�}�̒���2�`4mm�̂��̂��قƂ�ǁB�אڂ����3���}�Ԃ̋�����1.5�`3mm�ŁA�����̒Z�a�ȏ�ɊJ�����Ƃ͂Ȃ��B�̂�1�`1.2mm�قǂ̉~�`�A�e���}�̉����ł݂͌��ɗ���ĕ��z����B�������}��[�t�߂ł́A�ȉ~�`�A�p�����~�`�`���p�`�ɂȂ��Ė��ڂ��A�אڌ̂�䰕ǂ̈ꕔ�����L����B�u�ǂ̔��B�͈����A�ׂ��j���1���E2���u�ǂ��H�ɔF�߂�����x�B�����͔��B���邪�A�Ⴍ�ג����ȉ~�u��ŁA����ł͔F�����Â炢�B�����\�ʂɂ́A�ׂ����G���ȒP���[����ו���[�������B����B
�������F�T���S�ʈ�ł͕��ʂɌ����A���ɔg������̋����ʉ�����ʎΖʂ̐�(���[5m�Ȑ�)�ő���������B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{���Pocillopora eydouxi �w���W�J�n�i���T�C�T���S�ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Stylophora aff. Pistillata aff. pistillata (Esper, 1797)
�a���Ȃ� |
Pocilloporidae Gray, 1840 �n�i���T�C�T���S��
Stylophora Schweigger, 1819 �V���E�K�T���S�� |
|

�B�e�F�o�H���q(���V�\�s��v�ۍ`�A���[5m)
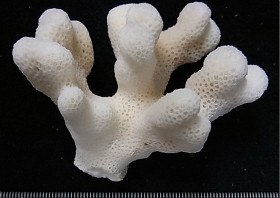
|

��1�E��2���}�̐�[�͞��`�`�~���`�B
|
|
�����`�F�w��`�R�����{�[�X��Q�́B�Q�̂͏��^�ŁA���a��20cm�����A���S�t�߂ł̍�����8cm�����̂��̂������B
��̕��̐F�ʂƓ����F��Ɋ��F�`�ΐF�A�S�̂������������������̂�����B
���i�̓����F��2���}�܂Ŕ��B����B��1���}�͑����ɐL�сA���̏㑤�ɏ������������2���}���قړ��Ԋu�Ŕz��B��1�E��2���}�̊T�ς͞��`�`�~���`�ŁA�����}�̊�̉��f�ʂ͉~�`�`�ȉ~�`�B��1���}�̒��a�܂��͒Z�a���P�`1.5mm�A��2���}�ł�5�`8mm�قǁB�}�̒����͑�1�E��2���}�Ƃ��ɒZ���A�O�҂̂قƂ�ǂ�5cm�����A��҂�2cm�����B�אڂ����2���}�Ԃ̋�����5mm�O��ŋ����B�T���S�͉̂~�`��䰌a��8mm�`1.2cm�A䰕ǂ����Ǐ�ɓˏo���邪�A䰕ǂ��}�̏�����Œ����� �ˏo������̂͏��Ȃ��B�����͑�2���}�̉����Ƒ�1���}��ł͂悭����Ă��邪�A��2���}�̐�[�߂��̃T���S�̂͊p�����~�`�ɂȂ�A�אڂ���T���S�̓��m��䰕ǂ̈ꕔ�����L���邱�Ƃ�����B䰓��ɂ͔��1���u�ǂ��悭���B���A䰐S�ɂ���Z���~����̎����Ɍ������ċ}�X�ŗ������ށB2���u�ǂ͔��B���Ă��Z��������ŁA䰕Ǐ�ł킸���ɔ��B���邩�����B�B�����\�ʂɂ͒P���[���`�ו���[�������B����B
�������F�ʉ�����ʎΖʏ㕔�A��ʎΖʂ̐��[10���Ȑ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Stylophora pistillata �V���E�K�T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A�����ł̐����ɂ��čČ������K�v�ł���BS. pistillata �́A�Q�̂͏����Ȏő���ŁA��1�E��2���}�̐�[�����`�ɂȂ邱�Ƃ͂Ȃ��B�܂��A���������ʒr���ʌΓ��ő���������B�����������R���痼��͕ʎ�Ǝv���A�X�Ȃ镪�ފw�I�������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FG. stokesi�Ƃ��ċL�^�����\������
|
|
| |
Goniopora cellulosa Veron, 1990
�n�`�m�X�n�i�K�T�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�����@�O(�������������哇�_�m�q�A���[2m)

�������ʐ^�Ɠ����Q�̂Ŏ�q���Y�ł͂Ȃ��B |

䰐S�̖_��ˋN���悭�ڗ��B
|
|
�����`�F��ɉ��Q�̂ŁA���ꂢ�ȃh�[����܂��͋���ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA�G��̐�[�ƌ��̕����͔����B���Ԃł��G��̒Z�������ȃ|���v��5mm�O��L�����Ƃ�����B
���i�̓����F�̂͊p�����~�`�A䰂͂��ꂢ�ȉ~�`�A䰂͒��a��2�`3mm�ŁA2mm�قǂ̌̂�����������B�܂��A���ΓI�ɌQ�̂̏���̌̂͏������A�����̂��̂͑傫���Ȃ�₷���B䰂̐[����2mm�����ŁA䰌a�̏������̂ł�䰂��[��������B䰕ǂ��k���Ō��ŁA�㉏�ɂ͐�[�̐�������������B���邽�߁A�g�Q�g�Q���Č�����B�u�ǂ�3���܂Ŕ��B���A�Z���_��̋��������ɕ��ԁB�������A2���u�ǂ͕����I�ɔ�ɂȂ邱�Ƃ�����B1���u�ǂ�䰂̉����ŁA2���u�ǂ͂����������ŐL�тāA�����܂ŒB����B3���u�ǂ́A䰕ǂ̓����ʂ���킸���ɒ���o�����x�̒����ŁA�����܂ŒB�����A�T�^�I�ȃn�i�K�T�T���S�^�̔z��p�^�[�����`�����Ȃ��B�����́A1���E2���u�ǂ̓����ɂł����傫��5�`6�̃p���A���̓����̏����Ȗ_��ˋN�Ƃ������Ȃ������̂Ŋ�{�I�Ɍ`�������B�������p�����������A������ēˏo�����_��ˋN�Ə����݂̂̂Ŏ������`�������̂̕��������B
�������F�ʒr�E��ʌ̐�Տ�A�g�Q�̉e���̏��Ȃ���ʎΖʂ̐�(����5m�Ȑ�)�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͎�q��(����q���剖��)�B�{���Goniopora burgosi �i�{�K�C�h���f�ڎ�j��G. tenuidens �}���n�i�K�T�T���S�Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��
|
|
| |
Goniopora djiboutiensis Vaughan, 1907
�L�N���n�i�K�T�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[8m)
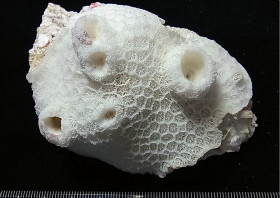
|

䰂̒��a��5mm�O��B
|
|
�����`�F�핢��`���Q�́B
��̕��̐F�ʂƓ����F�W���F�`�D�ΐF�≩�ΐF�ŁA���ƐG�肪�����ۂ��Ȃ�B���Ԃ��|���v�͂悭�L������B�G��͒Z���^�������A��[�͐�邱�Ƃ������B
���i�̓����F�̂�䰂����p�`�ŁA������䰂͒��a��5mm�O��A�H��6mm����̂�����Q�̓���1�`2�����邱�Ƃ�����B䰂͐[��1�`1.5mm���x�ƐA�u�ǂ͓ˏo�����A3���܂Ŕ��B�A�����͎����ɂ�炸�قړ����ŁA���Ԋu�ŋK���������z��B�u�ǂ̏㉏�Ƒ��ʂɂ́A��[�̐����������̋����Ɣ����ˋN���悭���B����B1���E2���u�ǂ�䰐S�߂��܂ŒB���ė��ݍ����A�X�|���W��̎������`������B�܂��A�u�NJԂɃV�i�v�e�B�L�����ւ��悭���B���邽�߁A�����|�̑啔���͎����Ɍ�����B3���u�ǂ�2���u�ǂ̔������x�̒����ŁA���̗���������ŗZ�����A�T�^�I�ȃn�i�K�T�T���S�^�̔z��p�^�[���ɂȂ�B1���E2���u�ǂ̓����ɂ́A5�`6�܂��͂���ȏ�̃p����䰐S���͂ނ悤�ɔ��B����B���̓����ɏ����Ȗ_��ˋN�������邱�Ƃ͋H�����A�����̂����B���邱�Ƃ�����B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʁA���p���ʌŌ�����B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Goniopora lobata �n�i�K�T�T���S�AG. cf.norfolkensis �I�I�n�i�K�T�T���S�AG. pendulus �����n�i�K�T�T���S�i�{�K�C�h���f�ڎ�j��G. aff.somaliensis �Ȃǂƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B�܂��A���i�`�Ԃ��悭�ގ�����G. cf.djiboutiensis �Ƃ̍X�Ȃ镪�ފw�I�������K�v�ł���B
* ��q�����L�^��
|
|
| |
Goniopora cf. djiboutiensis Vaughan, 1907
�a���Ȃ� |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[8m)

|

䰌a��5mm�O��B
|
|
�����`�F�핢��`���Q�́B
��̕��̐F�ʂƓ����F�W���F�ŁA���ՂƐG��̐�[�������B�G�肪�悭�L�т��|���v�ł́A���ՊO�����W���F�̓_���~�ʼn�����邱�Ƃ�A���̕������������������邱�Ƃ�����B���Ԃł��|���v��G�肪�悭�L������B�G���[�ׂ͍����B
���i�̓����F�̂͑��p�`�A䰂͉~�`�`�p�����~�`�ɂȂ�B䰌a��5mm�O��A䰕ǂ͂悭�ˏo���A䰂͐[��1�`2.5mm�قǁB�u�ǂ�3���܂Ŕ��B�B�u�ǂ̏㉏�Ƒ��ʂɂ́A��[�̐����������̋����Ɣ����ˋN���悭���B����B1���E2���u�ǂ�䰐S�߂��܂ŒB���ė��ݍ����A�X�|���W��̎������`������B�܂��A�אڂ���u�NJԂɃV�i�v�e�B�L�����ւ��悭���B���邽�߁A�����|�̑啔���������Ɍ�����B3���u�ǂƃp���̔��B���s�K���܂��͖����B�̌̂����������A�n�i�K�T�T���S�^�̊u�ǔz��p�^�[����������ɂ������Ƃ������B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʁA���p���ʌŌ�����B
�����ł̕��z�F�É������ɓ��E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{��́A�����̐F�ʁA䰂̐[���ƃp���̔��B�x���������AGoniopora djiboutiensis �L�N���n�i�K�T�T���S�ɂ悭����B����A���킩�ʎ킩�X�Ȃ镪�ފw�I�������K�v�ł���B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FG. somaliensis�Ƃ��ċL�^�����\������
|
|
| |
Goniopora lobata Milne Edwards and Haime, 1851
�n�i�K�T�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�쑺�b��(����q���剖���A���[2m)

|

䰂̒��a�͒ʏ�3�`4mm�B
|
|
�����`�F���Q�̂ŁA���ꂢ�ȃh�[���^�܂��͋��`�ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F�W���F�`�Ê��F�ŁA���Ƃ��̎����G��̐�[�͔����B���Ԃł��|���v��G��͂悭�L������B�G��͒Z�߂ŁA���ݒ[�B
���i�̓����F�̂͑��p�`�A䰂͑��p�`���p�����~�`�ɂȂ�B䰂̒��a�͒ʏ�3�`4mm�ŁA�H��5mm����̂�����Q�̓���1�`2������B䰂͐[��2mm�O��A䰌a���������Q�̂�䰕ǂ������Q�̂ł́A���ۂ���䰂��[��������B�u�ǂ�3���܂Ŕ��B���A䰕Ǐ㉏�ŕs�K���ɓˏo����B1���E2���u�ǂ́A䰕Ǐ㉏�Ƒ��ʂł͒���o�����A�u�Ǐ㉏�ɕs�K���ɕ��ԓݒ[�̖_���ŔF���ł�����x�B�����|�ł́A�����͒����j��(2���u�ǂ̈ꕔ�͔�)�ɂȂ���䰐S�܂ŒB����B�����݂͌��ɗ��ݍ����āA�V�i�v�e�B�L�����ւƂƂ��ɃX�|���W��̕s���ĂȎ������`������B3���u�ǂ͒Z���A�㉏�̋����݂̂��ˏo���ĒZ������Ɍ�����B���̂��ߖ{��ł́A�n�i�K�T�T���S�^�̊u�ǔz��͕s���ĂɂȂ�₷���B�Q�̎����̌̂ł́A1���E2���u�ǂ̓����Ƀp���A�_��ˋN�⏬���̂��`������A���ĂȎ������`������邱�Ƃ�����B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ̐A�ʒr�E��ʌ̊�Տ�Ō�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{���Goniopora djiboutiensis �L�N���n�i�K�T�T���S�ȂǑ�^�̂̓�������ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Goniopora cf. norfolkensis Veron and Pichon, 1982
�I�I�n�i�K�T�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�k��T�q(���V�\�s��Óc�`�A���[4m)

|

䰂̒��a��2�`4mm�B
|
|
�����`�F�핢��`���Q�́B
��̕��̐F�ʂƓ����F�W���F�`���F�B���Ղ͌��Ƃ��̎��肪�킸���ɔ��F�܂��͔����F�ŁA���̎����͔Z���F�Ńh�[�i�c��ɉ������B���Ԃł��|���v��G��͒����L�т�B�G��͐�ׂ�B
���i�̓����F�̂͑��p�`�A䰂͑��p�`���p�����~�`�B䰂͒��a2�`4mm�̂��̂������A䰂̐[����2.5mm�O��Ő[�����߁A䰕ǂ��ˏo���Č�����B�Q�̎����̌̂�䰕ǂ������A䰌a��5mm���邱�Ƃ����Ȃ��Ȃ��B�u�ǂ�3���܂Ŕ��B���A�㉏�ɂ͐�[����������A�܂��͝G���ɂȂ����������悭���B����B�������A䰌a�������Ȍ̂ł�3���u�ǂ��قƂ�ǔ��B�����A�j��̋��������������邱�Ƃ������B1���E2���u�ǂ́A䰕Ǐ㉏����䰐S�Ɍ������ċ}�p�x�ŌX����B�����̊u�Ǔ����́A��������̂悤�Ɍ݂��ɗ��ݍ������Ƃ͂Ȃ��A�킸���ɔ��B�����V�i�v�e�B�L�����ւŎ������`�������B1���E2���u�ǂ̓�����䰐S�Ƀp����_��ˋN�͌`������Ȃ��B3���u�ǂ͒Z���A����炪�悭���B����䰌a�̑傫�Ȍ̂������A�n�i�K�T�T���S�^�̔z��p�^�[���͌`������Ȃ��B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʁA���p���ʌ̊�Տ�ł�������B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͋H�B
�⑫�F�{���Goniopora djiboutiensis �L�N���n�i�K�T�T���S�ȂǑ�^�̂̓�������ƍ�������Ă��邽�߁A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��

|
|
| |
Goniopora aff. somaliensis Vaughan, 1907
�a���Ȃ� |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[8m)

|

䰌a��3�`4mm�B
|
|
�����`�F�핢��Q�́B
��̕��̐F�ʂƓ����F�W���F�ŁA���ՂƐG��̐�[�������B�G��͒��Ԃł������L�т邪�A�|���v�͒����L�тȂ��B�G�肪�k�ނƁA���Ղ��u��ɐ���オ���đ傫��������B�G��͐�ׂ�Ő�邪�A�F�ʂ��������ߓݒ[�Ɍ�����B
���i�̓����F�̂�䰂����p�`�`���ג������p�`�ŁA�Q�̎����̌̂͂��̎����������₷���B䰂͒��a��3�`4mm�A�H��5mm�ɒB����̂�����Q�̓���1�`2�����邱�Ƃ�����B䰕ǂ͂��܂�ˏo�����A䰂̐[����1�`2.5mm�قǁB�u�ǂ�3���܂Ŕ��B���A�����̏㉏�Ƒ��ʂɂ́A��[�̐����������̋����Ɣ����ˋN�������ɔ��B����B�܂��S�Ă̊u�ǂ�䰕Ǐ㉏�œ����x�ɔ������B1���E2���u�ǂ̑�����䰐S�܂ŒB���A�����̐�[��䰐S�߂��ŗ��ݍ����ăX�|���W��̎������`������B1���E2���u�ǂ̓����ɂ́A5�`6�̃p����䰐S���͂ނ悤�ɔ��B���邱�Ƃ�����B�܂��A�p���Ɉ͂܂��悤�ɁA䰐S�ɏ����̂Ə����Ȗ_��ˋN�������邱�Ƃ�����B1���E2���u�ǂɔ�ׂ�ƁA3���u�ǂ͒Z���A䰐S�܂ŒB���Ȃ��B�悭���B����3���u�ǂ́A2������2���u�ǂ𗼑����狲�ݍ��ނ悤�ɐL�����A�T�^�I�ȃn�i�K�^�T���S�^�̊u�ǔz��p�^�[���ɂȂ�B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʁA���p���ʌŌ�����B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Goniopora somaliensis �\�}���A�n�i�K�T�T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Goniopora tenuidens (Quelch, 1886)
�}���A�i�n�i�K�T�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Goniopora de Blainville, 1830 �n�i�K�T�T���S�� |
|

�B�e�F�[���T�L(����q���剖���A���[�s��)

|

䰌a��2�`3.5mm�B
|
|
�����`�F��ɉ��Q�̂ŁA���ꂢ�ȃh�[���^�܂��͋��`�ɂȂ邱�Ƃ������B
��̕��̐F�ʂƓ����F��ɒW���F�ŁA���ΐF��D���F�Ȃǖ��邢�F�ʂɂȂ邱�Ƃ��������A���Ƃ��̎��肪�킸���ɔ����B�G��͒Z�߂ŁA���������[�܂ő������������A��[�͐�ׂ�ɂȂ炸�ۂ��B
���i�̓����F�̂͊p�����~�`�A䰂͂��ꂢ�ȉ~�`�ɂȂ�B䰌a2�`3.5mm�A䰂̐[����1.5mm�O��B�u�ǂ�3���܂Ŕ��B�B�u�Ǐ㉏�ɂ�������܂��͒Z���_��̋��������B���A�قړ��Ԋu�Ŕz��B1���E2���u�ǂ́A�u�ǂ̑��ʂł͔��ɒZ���A������̋����̑��݂ɂ���Đh�����ĔF���ł�����̂�����B�����|�ł́A�����͒�������(2���u�ǂ̈ꕔ�͔�)�ɂȂ�A䰐S�߂��܂ŒB����B䰐S�߂��ł�1���E2���u�ǂ̊ԂɃV�i�v�e�B�L�����ւ��A�����ɂ�5�`6�̃p�����A�����̓����ɂ͏����̂Ɩ_��ˋN�����B���A�k���Ō��łȎ������`������B����2���u�Ǔ����̃p���͂悭������ď���ɓˏo����B
�������F�g�Q�̉e���̏��Ȃ��ʎΖʂ��ʎΖʂ̐�A�ʒr�E��ʌ̊�Տ�Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
�⑫�F�{��͐G��̌`���̂̑傫�����ގ�����Goniopora lobata �n�i�K�T�T���S�Ȃǂƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B
* ��q�����L�^��
|
|
| |
Porites australiensis Vaughan, 1918
�n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|

�B�e�F���n�m�V(���V�\�s���Y�A���[10m)
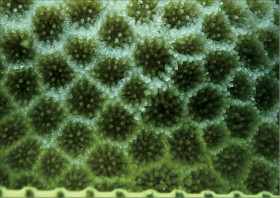
|

䰌a��1.5mm���x�B
|
|
�����`�F���a��m�̋���ȉ��Q�̂ɂȂ�B�Q�̕\�ʂ͊��炩�ŁA�召�l�X�ȕs�K����ᎂ�������B
��̕��̐F�ʂƓ����F�N���[���F�A�݂������������F�ȂǁB
���i�̓����F�̂�䰌a��1.5mm���x�A���ɂ�2mm�ɒB������̂�����B䰕ǂ͔�r�I�����A�אڂ���̊Ԃ�������ɂȂ�B�u�Ǐ�ɂ͒ʏ�2�̎���ˋN���F�߂��A䰕ǂƑ��܂��Č̑S�̂ɓ��S�~��̍\���������ł���B�O���͒ʏ�V�����邪�A�V�i�v�e�B�L�����ւŎO����Ɍq���邱�Ƃ�����B�悭���B����8�{�̃p��������A���̂������u�Ǒ̍��E2��4�{�͓��ɑ傫���A�����ł��ڗ��B�����͒���܂��ׂ͍���ł���B
�������F�ʒr����ʎΖʂ܂ŕ��ʂɌ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͏��Ȃ��B
* ��q�����L�^��
|
|
| |
Porites heronensis Veron, 1985
�t�^�}�^�n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|

�B�e�F���n�m�V(���V�\�s�Y�c�p�A���[7m)

|

䰌a��1�`1.5mm���x�B
|
|
�����`�F�\�ʂ͊��炩�ŁA�s�K����ᎏ�̐���オ������핢��Q�̂������B���ɂ�ᎂ⒌������^�̉��Q�̂�������B
��̕��̐F�ʂƓ����F�N���[���F�A���F�A�݂��������W�D�F�ȂǕψقɕx�ށB
���i�̓����F�̂�䰌a��1�`1.5mm���x�ŁA䰂͐B�̂̊Ԋu�͗l�X�ŁA�L���Ƃ���ł�1mm����B�O���͎O����ɂȂ邪�A�u�ǔz�s�K���ŁA���u�ǑƂ��F�߂������̂������B�p����7�{�ȉ��ŁA�u�ǐ�[���q���~�`�̃V�i�v�e�B�L�����ւ��ǂ����B����B�����͍Y��ł��邪�A�������܂ތ̒����̍��i�\�������S�Ɍ����A�������悤�Ɍ�����̂������B
�������F���p��`�̒��Ȃǂ̎Օ��I�Ȋ�����A�g�Q�̉e�����傫����ʈ�̐��܂ōL����������B
�����ł̕��z�F��t���َR�E���茧�Δn�`����{���B��q�����܂ޒg���ш�ł͕��ʎ�ŁA�n�}�T���S���̒��ł͍ł��k�܂Ō������ł���B
* ��q�����L�^��
|
|
| |
Porites lobata Dana, 1846
�t�J�A�i�n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|

�B�e�F���n�m�V(���V�\�s�Y�c�p�A���[6m)
|

䰌a��1.5mm�ȏ�B
|
|
�����`�F���a��m�̋���ȉ��Q�̂ɂȂ�B�Q�̕\�ʂɂ͑召�l�X�ȕs�K����ᎂ�������B
��̕��̐F�ʂƓ����F�Ê��F�≩���F�ȂǁA�ψقɕx�ށB
���i�̓����F�̂�䰌a��1.5mm���x����������傫���A䰕ǂ͔����A�O�`�͑��p�`��悷��B�O���͏�ɗV�����A�p���̔��B�͎ア�BPorites solida�I�I�n�}�T���S�Ɏ��邪�A�{��̕������䰂����������ƁA8�{�̃p���Ƃ悭���B������̎����������ƁA䰌a�ɑ���u�ǂ̐�[�ň͂܂�钆���|�̔䗦���ꌩ���ď��������Ƃŋ�ʂł���B
�������F�ʒr����ʎΖʂ܂ōL��������B
�����ł̕��z�F���m�������ȓ�B��q���ł͏��Ȃ��B
* ��q�����L�^��
|
|
| |
Porites lutea Milne Edwards & Haime, 1860
�R�u�n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|

�B�e�F���n�m�V(���V�\�s��v�ۍ`�A���[4m)

|

䰌a��1.5mm���x�B
|
|
�����`�F���a��m�̋���ȉ��Q�̂ɂȂ�B�Q�̕\�ʂ͊��炩�ŁA�召�l�X�ȕs�K����ᎂ�������B
��̕��̐F�ʂƓ����F�����F��݂����������F�ȂǁA�ψقɕx�ށB
���i�̓����F�̂�䰌a��1.5mm���x�ŁA䰕ǂ͔�r�I�����B�O���͂قڗ�O�Ȃ��Z�����邩�O����Ɍq����B�p����5�`6�{�A����8�{�܂ŔF�߂��A���̂������u�Ǒ�4�{�ƎO����1�{�̌v5�{�����ɖڗ����A�����ɂ����ĂɔF�߂��邱�Ƃ������B�����͂悭���B���A����܂��͝G����������ŁA�����̂ƌĂ����ˏ�̍\���Ŋu�ǂƌq����BPorites australiensis�n�}�T���S�ɍ������邪�A䰕ǂ���┖���O�����Z�����邱�ƁA5�{�̃p�����ڗ����ƂŁA���낤���ċ�ʂł���B
�������F�ʒr����ʎΖʂ܂ōL��������B
�����ł̕��z�F�a�̎R�����{�E�F�{���V�����[�ȓ�B��q���ł͂�╁�ʁB
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FP. superficialis�Ƃ��ċL�^�����\������

|
|
| |
Porites okinawensis Veron, 1990
�I�L�i���n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|

�B�e�F���n�m�V(���V�\�s��v�ۍ`�A���[3m)

|
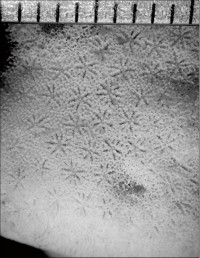
䰌a��1.2�`1.7mm�B
|
|
�����`�F���^�̔핢��܂��͉��̌Q�̂ŁA�\�ʂ͊��炩�ł���B
��̕��̐F�ʂƓ����F���F����W���F�B
���i�̓����F�̂�䰌a1.2�`1.7mm�ŁA�قƂ�lj��܂����ʓI�ł���B�̂̔z��͕s�K���ŁA�אڌ̂���������Ƃ��������ΊԊu��2mm�قǂɂȂ�Ƃ��������B�O���͏�ɗZ�����邪�A�u�ǂ̖����͌̂̑傫���ɂ���ĕω����A�n�}�T���S���ɓ����I�Ȕz������ɂ������Ƃ�����B�u�ǂ͒��S�t�߂ɒB���A�ꕔ�͎����ƗZ������B�p���̔��B�͎キ�A�S�����@����̂������B�����̓����͐����ł��A�|���v���L�тĂ��Ȃ��Q�̂ł͖��ĂɔF�߂��邽�߁A�ꌩ���Ė{��ƕ�����B
�������F���p��`�̒��ȂǎՕ��I�Ȋ��Ɍ����邪�A���܂葽���Ȃ��B
�����ł̕��z�F��t���َR�`���\���Ŋm�F����Ă�����{�̌ŗL��ŁA�n�}�T���S���̒��ł͍ł����z�悪�L���B��q���ł͋H�B
�⑫�F�{��̃^�C�v�Y�n�͉���{�������p�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Porites solida (Forskål, 1775)
�I�I�n�}�T���S |
Poritidae Gray, 1840 �n�}�T���S��
Porites Link, 1807 �n�}�T���S�� |
|
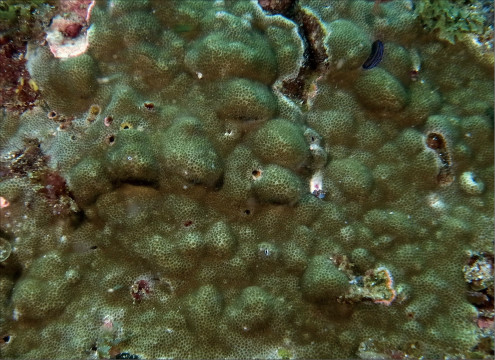
�B�e�F���n�m�V(���V�\�s�匴�A���[8m)

|

䰌a��2mm�O��Ƒ傫���B
|
|
�����`�F��^�ɂȂ���Q�̂ŁA�\�ʂ͔g�ł��召�l�X��ᎂ�����B
��̕��̐F�ʂƓ����F�������������F�B
���i�̓����F�{��͓̌̂��{�Y�n�}�T���S���ł͍ł��傫���A���ɌQ�̂̓ʕ��ł�䰌a2mm���邱�Ƃ�����B�͔̂����č���䰕ǂŎd��ꂽ���p�`�ŁA䰂��[���A�Z���I�C�h�^�ɔz��B�����̓�������APorites lobata�t�J�A�i�n�}�T���S�ȊO�̎�Ƃ̋�ʂ͗e�Ղł���B�O���͗V�����A�p���͂Ȃ����A�����Ă����B���キ�{�������Ȃ��B�����͝G���ŒႢ����Ȃ��A���������܂钆���|���t�J�A�i�n�}�T���S�ɔ�ׂčL���B
�������F�ʒr��ʎΖʂ̐Ɍ����邪�A���܂葽���Ȃ��B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
* ��q�����L�^��
|
|
| |
Psammocora albopicta Benzoni, 2006
�x���x�b�g�T���S |
Psammocoridae Chevalier and Beauvais, 1987 �A�~���T���S��
Psammocora Dana, 1846 �A�~���T���S�� |
|

�B�e�F�[���T�L(����q���n���̊≮�A���[5m)

|
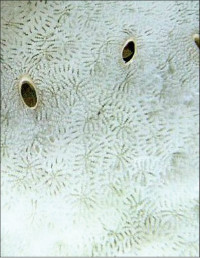
1�`1.5mm�Ə������B
|
|
�����`�F�핢��Q�́B��Ղ�핢���Ȃ���A���a��50cm�ȏ�̑�^�ɂȂ邱�Ƃ������B�Q�̂ɂ���ẮA�\�ʂɒ��a2�`3cm�A����1cm���x�̉~�u��̕s�K���Ȑ���オ�肪�����`�����ꂽ��A��������Ղ���V�����āA���t��`��ɒ���o�����肷�邱�Ƃ�����B
��̕��̐F�ʂƓ����F��ɐԊ��F�B���ш�ł͑N�₩�ȉ��ΐF�ɂȂ�A���Ԃł��Z���G���L�����߁A�Q�̕\�ʂ��r���[�h��̌���������Ƃ������B�̂̌��Ղ������W���F�ɂȂ邱�Ƃ�����B
���i�̓����F�̂͑��p�`�A䰂̒��a��1�`1.5mm�ŏ��^�A�T���i�X�e���C�h�^�z��B䰂͐A�̊Ԋu��1mm�����ŋ������߁A�Q�̕\�ʂ������Ɍ�����B�\�ʂɉ~�u��̓ˋN�����Q�̂ł́A�Z���w���Ⴂ�R�������`������邱�Ƃ�����B���̏ꍇ�A䰂��[���A�̔z�Z���I�C�h�^�܂��͂��A���h���C�h�^�Ɍ�����B�u�ǁE�]��3���܂Ŕ��B�B1���E2���u�ǂ͂قړ��������ŕ��ˏ�ɔz�A䰐S�Ɍ������ɂ�Ĕ����Ȃ�A��M��̏����Ȏ����ɒB����B3���u�ǂ͒Z���A1���E2���u�ǂ̔����ȉ��̒����B�Q�̂ɂ���ẮA1���E2���u�ǂ������I�ɗZ�����ĉԕُ�̔z��ɂȂ邱�Ƃ�����B�܂��A�אڂ���u�ǁE�]�̊Ԃ��V�i�v�e�B�L�������w偂̑���ɂȂ��A�V�i�v�e�B�L�����ւ��`������邱�Ƃ�����B�u�ǁE�]�̏㉏�ɂ́A��[���������ɕ����������悭���B���邽�߁A�u�ǁE�]�͎��ۂ��������A�Q�̕\�ʂ̓U���U�����Č�����B
�������F�ʎΖʂ܂��͊�ʎΖʂ̐��[15m�Ȑ[�Ō�����B�J���I�Șp���Ȃǂ̔g������̎ア�Ƃ���ł́A�����[�Ō����邱�Ƃ�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͋H�B
�⑫�F�{���Psammocora profundacella �A�~���T���S��P. nierstraszi �q�_�A�~���T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A��q���ȓ�ł̐����ɂ��Ă͍Č������K�v�ł���B�x���x�b�g�T���S�̘a����S���̂�P. superficialis �ł͂Ȃ��{��ł���B
�����EVeron (1995)�ł̏o���L�^�F���iB. wellsi�Ƃ��ċL�^�j
|
|
| |
Psammocora profundacella Gardiner, 1898
�A�~���T���S |
Psammocoridae Chevalier and Beauvais, 1987 �A�~���T���S��
Psammocora Dana, 1846 �A�~���T���S�� |
|

�B�e�F�[���T�L(����q���n���̊≮�A���[5m)
|
䰂̒��a��2�`5mm�B
|
|
�����`�F�핢��`�����Q�́B�T���S�ʈ�ł͒��a10cm�����̂��̂��������A���т̊�ʈ�ł�30cm�ȏ�ɂȂ���̂��������Ȃ��B�Q�̎�������Ղ���V�����邱�Ƃ͂Ȃ��B
��̕��̐F�ʂƓ����F��ɗΐF�`���F�ŁA���ΐF�Ȃǂ̖��邢�F�ʂɂȂ���̂�����B�핢��Q�̂ł́A�̂̌��Ղ������W���F�ɂȂ邱�Ƃ�����B���ш�ł́A���Ԃł����ڂŐ�ׂ�̒Z���G����悭�L���B�Q�̕\�ʂ̓U��������ۂ���B
���i�̓����F�̂͑��p�`�A�傫���͕s������䰂̒��a��2�`5mm�B�ʏ�A�̔z��̓T���i�X�e���C�h�^�����A�אڂ���̊Ԃɏ㉏�̔����R����(�H�Ƀ����e�B�N��)�����B���邽�߁A����ł̓Z���I�C�h�^�܂��̓��A���h���C�h�^�Ɍ�����B䰓��ɂ�1�`3�̓��S�~��̃V�i�v�e�B�L�����ւ��悭���B����B䰂͐A3���u�ǁE�]�܂Ŕ��B����B�����̊u�ǂ̌����ƍ����͂قړ����ŁA1���E2���u�ǂ͒����A䰐S�܂ŒB����B3���u�ǂ͑��̊u�ǂ̔����ȉ��̒����ŒZ���B�Q�̂ɂ���ẮA1���E2���u�ǂ̔z�y�^���C�h�^�ɂȂ邱�Ƃ�����B�u�ǁE�]�̏㉏�ɂ́A��[���������ɕ����������悭���B����B���̂��߁A�u�ǁE�]�͌����A�אڂ���u�ǁE�]�Ԃ͋����A�Q�̕\�ʂ̓U���U�����Č�����B䰐S�ł́A1���E2���u�ǂ̓�������L�т��g���x�L���������ݍ����B���̏㕔�ɂ́A�ݒ[�ŒZ���_���ƁA�������͂ނ悤�ɕ����̒Z���p�����`�������B�����͎��͂̊u�Ǔ��������Ⴍ�A䰐S�ɉ~�`�̒����|���F�߂���B
�������F�ʒr�E��ʌΓ���A�g������̏��Ȃ��ʎΖʂ��ʎΖʂŌ�����B
�����ł̕��z�F��t���َR�E�������B���ȓ�B��q���ł͋H�B
�⑫�F�{���Psammocora albopicta �x���x�b�g�T���S��P. nierstraszi �q�_�A�~���T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă��邽�߁A��q���ȓ�ł̐����ɂ��Ă͍Č������K�v�ł���B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Blastomussa merleti (Wells, 1961)
�J�r���^�o�T���S |
Incertae sedis �����Ȗ���
Blastomussa Wells, 1968 �I�I�^�o�T���S�� |
|

�B�e�F�[���T�L(�B�e�n�_�Ƃ��̐��[�A�W�{�̏��݂͕s��)
|

䰌a��4�`6mm�قǁB
|
|
�����`�F�ג����~����̌̂������W�܂�������Q�́B
��̕��̐F�ʂƓ����F���F�`�ΐF�B���Ղ͎��͂������邢�F�ʂɂȂ邱�Ƃ������B�|���v�ƐG��͒��Ԃł��킸���ɐL�т�B�|���v�\�ʂł́A���A��ɖc����܂肪���ˏ�ɔz��B
���i�̓����F�̂͂��ꂢ�ȉ~�`�ŁA䰌a��4�`6mm�قǁA�̂̑傫����אڂ���̊Ԃ̋����͂悭�����B�̔z��͎�Ƀt�@�Z���C�h�^�����A�����I�ɗאڌ̂��Z�����A���v���R�C�h�^�`�Z���I�C�h�^�ɂȂ邱�Ƃ�����B�u�ǂƘ]��3���܂Ŕ��B�B1���E2���u�ǂ̒���������͂قړ����ŁA���ꂢ�ȕ��ˏ�ɔz�A�����܂ŒB����B�u�ǂ�䰕Ǐ㉏�܂��͂��̓����Ō����A1�`2mm�قǓˏo���A��������䰐S�Ɍ������ċ}�X�ŗ������݂Ȃ��甖���Ȃ�B3���u�ǂ͔��ɔ����Z���A䰕Ǐ㉏�`�����ł킸���ɔ��B������x�B�u�Ǐ㉏�ɂ͋��������B���邪�A�������Ėڗ����Ȃ��B�u�Ǒ��ʂɂ�������ˋN�����B�B1���E2���]�͌��������ɒZ���B3���]�͒ʏ햢���B�ŁA��^�̂ŋH�ɔ��B���邱�Ƃ�����B�����͉~�`�ŁA䰐S�ɂ��鏬���Ȗ_��(�H�ɔ�)�̓ˋN�A���̓ˋN��1���E2���u�Ǔ������Ȃ����ˏ�̏����́A�אڂ���u�Ǔ������Ȃ���̃g���x�L�����Ō`�������B�̂̊O���\�ʂɂ̓G�s�e�J���悭���B����B�܂�䰕Ǔ����ɁA�����ȖA���g�D���`������邱�Ƃ�����B
�������F�ʎΖʂ̐[���g�Q�̉e�������Ȃ���ʎΖʂ̂悤�ȁA�������������D�����܂�₷�����Ō�����B
�����ł̕��z�F��q���ȓ�B��q���ł͋H�B
* ��q�����L�^��
�����EVeron (1995)�ł̏o���L�^�FL. pruinosa�Ƃ��ċL�^�����\������

�������ʐ^�̌Q�̂Ƃ͕ʌQ��(���ꌧ���\���Ԏ�p�A���[27m�ō̏W)
|
|
| |
Blastomussa vivida Benzoni, Arrigoni and Hoeksema, 2014
�I�I�^�o�T���S |
Incertae sedis �����Ȗ���
Blastomussa Wells, 1968 �I�I�^�o�T���S�� |
|

�B�e�F�쑺�b��(����q���n���≮�A���[10m)
|
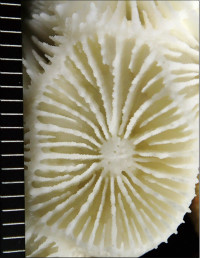
䰂̒��a��1.2�`2.5cm�ŕs�����B
|
|
�����`�F�w�̒Ⴂ����Q�́A�����`���Q�́B
��̕��̐F�ʂƓ����F��ɊD���F��D�ΐF�B�Ԃ݂�тт����̂�������B�|���v�͒��Ԃł��O������ɂ悭�L�сA�\�ʂɐ��A��ɖc����܂肪���ˏ�ɔz�邱�Ƃ�����B�G��́A�O�������̎����ƁA���A���̓����߂�����킸���ɐL�т�B�|���v�����ƐG���[�́A���̕�����蔒���ۂ�������B
���i�̓����F�̔z��̓t�@�Z���C�h�^�A���v���R�C�h�^�`�Z���I�C�h�^�B�t�@�Z���C�h�^�⏀�v���R�C�h�^�͉̌̂~�`�`�ȉ~�`�A�Z���I�C�h�^�̌̂͊p�����~�`�`���p�`�B�̂̒��a��1.2�`2.5cm�قǁA�傫���͕s�����B�u�ǁE�]�͒ʏ�4���܂ŁA䰌a�̑傫�Ȍ̂ł�5���܂Ŕ��B���邱�Ƃ�����B�u�ǁE�]��������ˋN���悭���B����B1���E2���u�ǂ͌����A������ɓˏo����B1�`3���u�ǂ͎����܂ŒB����B4���u�ǂ͔����A�����Ă������̎�O�܂ŁB5���u�ǂ͂���ɔ����Z���A䰕Ǐ㉏�ł킸���ɔ��B����̂݁B��[���ۂ݂�тт����[�u��̋������A�S�Ă̊u�Ǐ㉏�ɂ悭���B����B䰕Ǐ㉏�����������̋������ł��傫���悭�ˏo����B�܂��A�u�Ǐ㉏��䰐S�Ɍ������ĂقƂ�njX���Ȃ����߁A䰂͂��Ȃ������B�]��䰕NJO���ł킸���ɐ���オ����x�ŁA�������`������邱�Ƃ͋H�B�����͉~�`�`�ȉ~�`�ŁA䰐S�̖_��(�H�ɔ�)�ˋN�A���̓ˋN��1�`3���u�Ǔ����̊Ԃɂł��鑽���̃p���A���҂��Ȃ������́A�u�Ǔ�����p���Ԃ��Ȃ��g���x�L�����Ō`�������B
�������F�g�Q�̉e�������Ȃ��ʎΖʂ��ʎΖʂ̐��[10m�Ȑ[�Ō�����B�Օ��I�ȓ��p�ł͂��Ƃ���ł�������B
�����ł̕��z�F�a�̎R�����{�E���茧��ܓ����ʓ��`��q���B��q���ł͋H�B
�⑫�F�{��͂���܂�Blastomussa wellsi �i�{�K�C�h���f�ڎ�j�ƍ�������Ă����B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Leptastrea bewickensis Veron, Pichon and Wijsman-Best, 1977
�q�������T���S |
Incertae sedis �����Ȗ���
Leptastrea Milne Edwards and Haime, 1848 �����T���S�� |
|

�B�e�F�[���T�L(���V�\�s���Y�A���[12m)

|

䰂͒��a4�`5mm�B
|
|
�����`�F�핢��Q�́B�Q�̕\�ʂ͕��R�ŁA�R�u��ɐ���オ�邱�Ƃ͏��Ȃ��B
��̕��̐F�ʂƓ����F��Ɋ��F�B�Q�̕\�ʂ̐F�ʂ��܂���ɂȂ���̂�������B�̊Ԋu�͋����A�אڌ̂Ƃ̋��E�͕s���āB���Ԃł��G����悭�L�����A�悭�ˏo����1���u�ǂ͔F���ł���B
���i�̓����F�̂͑��p�`�ŁA�Z���I�C�h�^�ɔz��B䰂̒��a��4�`5mm�̌̂��قƂ�ǂŁA�Q�̏�ł̌̂̑傫����`�͂悭�����B�u�ǂ�3���܂Ŕ��B���A䰕Ǐ㉏�œˏo����B����1���u�ǂ��悭�ˏo���邽�߁A�Q�̕\�ʂ͑e��������B1���u�ǂ͒����A䰐S�܂ŒB����B2���u�ǂ͒Z���A�������̂ł�1���u�ǂ�1/2�قǁB3���u�ǂ͂���ɒZ���A䰕Ǐ㉏�ł킸���ɔ��B������x�B�����́A䰐S�ɂ���1�{�̏����Ȗ_��ˋN�A���̊O����1���E2���u�ǂ̓����ɔ��B����2�`3�̃p���A���҂�A�����鏬���̂���Ȃ邪�A�����̂ǂꂩ�������B�܂��͕s���ĂŁA�������ڗ����Ȃ����Ƃ������B
�������F�ʒr���ʌ̊�Ղ��L���I�o����Ƃ����A�ʎΖʁE��ʎΖʂŌ�����B
�����ł̕��z�F��q���ȓ�B��q���ł͕��ʎ�B
�����EVeron (1995)�ł̏o���L�^�F��

|
|
| |
Leptastrea aff. pruinosa Crossland, 1952
�g�Q�����T���S |
Incertae sedis �����Ȗ���
Leptastrea Milne Edwards and Haime, 1848 �����T���S�� |
|

�B�e�F���n�m�V�i����q���n���̊≮�A���[10m�j

|

䰂̒��a��3mm�`1cm�ŕs�����B
|
|
�����`�F�핢��`�����Q�́B�Q�̕\�ʂ��s�K���ɃR�u��ɐ���オ�邱�Ƃ������B��ʈ�̓��p�ł́A�C��ɓ]�������I���Ȃ��琬�����邱�Ƃ�����B
��̕��̐F�ʂƓ����F�ΐF�`�Ԋ��F�B���Ԃł��ݒ[�ȐG���L�����߁A䰕Ǔ���������オ���Č�����B�܂��A����ɔ����אڌ̂̋��E�ɍa���������悤�Ɍ�����B
���i�̓����F�͎̂�ɑ��p�`�ŁA�v���R�C�h�^�܂��̓Z���I�C�h�^�ɔz��B䰂̒��a��3mm�ɖ����Ȃ��̂���1cm����̂܂ŗl�X�ŁA�`��傫�����s�����Ɍ����₷���B�u�ǂ͒ʏ�4���܂Ŕ��B���A���ʂɒZ������ˋN���悭���B����B���̂��ߊu�ǂ͌����A�אڂ���u�NJԂ�����������B�܂��A䰕Ǐ㉏�ł�1���E2���u�ǂ�3���E4���u�ǂ̌�����ˏo�x���͂قړ����Ȃ̂ŁA�e�̂��ˏo���Č����邱�Ƃ͂Ȃ��B1���E2���u�ǂ͒����A䰐S�̎����܂ŒB����B�ʏ�3���u�ǂ�1���E2���u�ǂ����Z�����A�����߂��܂Ŕ��B���邱�Ƃ�����B���̏ꍇ�A3���u�ǂ̓������אڂ���2���u�ǂ̑��ʂɗZ�����āA�u�ǂ̔z�s�K���Ɍ�����B4���u�ǂ͔��ɒZ���A�L�тĂ�3���u�ǂ�1/2���x�̒����B3���E4���u�ǂɔ�ׂ�ƁA1���E2���u�ǂ�䰕Ǐ㉏�̂���������䰐S���ɂ��ˏo����B�����́A䰐S�ɂ��鏬���Ȗ_��ˋN�A���̊O����1���E2���u�Ǔ����ɔ��B���������̃p���Ə����̂���Ȃ�B�p����3���u�ǂ̓����ɂ��`������邱�Ƃ�����A���̏ꍇ�͎��������ɑ傫���Ȃ�B
�������F��ʎΖʁA�ʎΖʂ��ʌ̊�Ղ��L���I�o����Ƃ����A��ʌ���p�̍��I����I��Ō�����B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���ł͕��ʎ�B
�⑫�F�{���Leptastrea pruinosa ��L. purpurea �����T���S�i�{�K�C�h���f�ڎ�j�ƍ�������Ă���A�����ł̐����ɂ��Ă͍Č������K�v�ł���B

|
|
| |
Oulastrea crispata (Lamarck, 1816)
�L�N���C�V���h�L |
Incertae sedis �����Ȗ���
Oulastrea Milne Edwards and Haime, 1848 �L�N���C�V���h�L�� |
|

�B�e�F�쑺�b��(����q���n���̊≮�A���[12m)
|

䰂̒��a��4�`5mm�B
|
|
�����`�F���^�̔핢��`�����Q�̂ŁA�Q�̂̒��a��10cm���邱�Ƃ͖ő��ɂȂ��B
��̕��̐F�ʂƓ����F���F�`�Ê��F�B��u�ǁE�]�̏㉏�������ۂ������A�悭�ڗ��B���Ԃł��G���L�����Ƃ�����B
���i�̓����F�u�ǂƘ]�������������̍��i�����F�܂��͊��F��悷��B�̂͊p�����~�`�����p�`�ŁA䰂̒��a��5�`7mm�B�̔z��͎�Ƀv���R�C�h�^�����A�̊Ԋu�������Q�̂ł̓Z���I�C�h�^�ɁA�L���Q�̂ł̓T���i�X�e���C�h�^�Ɍ����₷���B�u�ǂƘ]��4���܂Ŕ��B���A��̂��̂قnj����X��������B�u�ǂƘ]�̑��ʂ���㉏�ɂ����ẮA��[�̐����������ˋN���悭���B����B���ɑ��ʂł́A����炪�����̗���Ȃ��B1���E2���u�ǂ͎����܂ŒB���A�����̂����O���Ƀp����t�����B����B�����́A�p����t�̓����Ɍ`�����ꂽ�p���������W�܂��Č`�������B����������ł̃p����t�ƃp���̎��ʂ͍���B�̕\�ʂ���͔F�����Â炢���A䰕ǂ͔�ŁA�����̏���͑��E���B�Q�̗��ʂɂ͖��ĂȃG�s�e�J�͔F�߂��Ȃ��B
�������F���D�����܂�₷�����p�̐�A�g�Q�̉e���̏��Ȃ���ʎΖʂ̐��[10m�Ȑ[�Ō�����B
�����ł̕��z�F��t�����Y�E�V�������n���ȓ�B��q���ł͋H�B

|
|
| |
Plesiastrea versipora (Lamarck, 1816)
�R�}���L�N���C�V |
Incertae sedis �����Ȗ���
Plesiastrea Milne Edwards and Haime, 1848 �R�}���L�N���C�V�� |
|

�B�e�F�i�c�r��(���V�\�s���Y�A���[10m)

|

䰌a�͒ʏ�3�`4mm�B
|
|
�����`�F�핢��`�����Q�́B
��̕��̐F�ʂƓ����F>��Ɋ��F�ŁA���ΐF��D�ΐF�̂��̂�����B�g���ш�ł́A���Ԃł��G���L�����Ƃ�����B�אڌ̂Ƃ̋��E�ɒ�����̂��ڂ݂�����B
���i�̓����F�͎̂�ɉ~�`�ŁA䰌a�͒ʏ�3�`4mm�B�̔z��̓v���R�C�h�^�ŁA�̊Ԋu�͋����B�u�ǁE�]��3���܂Ŕ��B����B1���E2���u�ǂ͌����A䰐S�܂ŒB���邪�A3���u�ǂ͒Z���A䰐S�ɒB���邱�Ƃ͂Ȃ��B1���E2���u�ǂ̓����ɂ́A������č����ˏo�����p�������B���A����炪������ɔz��B���̓����ɂ́A�������ĒZ�������̖_��ˋN����Ȃ鎲����������B�Q�̎����̌̂������A�]�͒Z�߂ŁA�אڂ���̊Ԃŋ��L����邱�Ƃ͂Ȃ��B䰕ǂ͌����A�����\�ʂ���킸���ɓˏo���邽�߁A�̂͊�ɐ���オ���Č�����B�Q�̗��ʂɖ��ĂȃG�s�e�J�͌`������Ȃ��B
�������F�ʏ�A����{���ȓ�ł͏ʎΖʂ̐��[20m�Ȑ[�Ō�����B����{���Ȗk�ł͏ʒr����Օ��I�ȓ��p�̐ł�������B
�����ł̕��z�F��t���َR�E���茧�Δn�ȓ�B��q���Ȗk�ł͕��ʎ�B

|
|
<< �߂��@�@�i1/1)
�����₢���킹�F�@�������������@�������l���̈�
��305-8506 ��錧���Ύs�����16-2�@Email: biodiv.data@�i���̕����͍폜���ă��[�����ĉ������jnies.go.jp

