The World of Protozoa, Rotifera, Nematoda and Oligochaeta
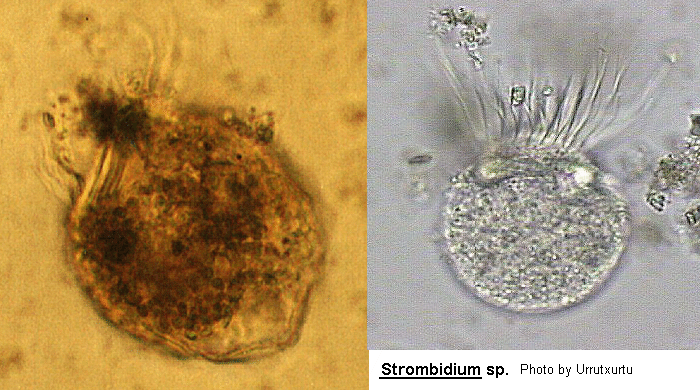
Strombidium
Strombidium Claparede & Lachmann, 1858 (ref. ID; 3388, 3420), 1859 (ref. ID; 3690, 4906) or 1861 (ref. ID; 2014)
From Dr. Inaki
Class Polyhymenophora: Subclass Spirotricha: Order Oligotrichida: Suborder Oligotrichina (ref. ID; 2014)
Class Polyhymenophora Jankowski, 1967: Order Oligotrichida Butschli, 1887 (ref. ID; 4893)
Class Spirotrichea: Subclass Oligotrichia: Order Oligotrichida: Family Strombidiidae (ref. ID; 7456)
Strombidiida (ref. ID; 4849)
Family Strombidiidae Faure-Fremiet, 1970 (ref. ID; 4356) or 1979 (ref. ID; 4906)
[ref. ID; 2014]
Body shape an elongate ovoid or pyriform with the posterior body half encased in a lorica-like arrangement of closely fitting polysaccharide hexagonal platelets. The anterior half of the body cannot retract into the lorica. At the apical region there is a slight protuberance around which the conspicuous open AZM winds. Somatic cilia absent. Internally there is a series of oblique trichites with a skeletal function.
Most species of the genus are marine but there are a few freshwater representatives.
Quote; Colin R. Curds, Michael A. Gates and David McL. Roberts "British and other freshwater ciliated protozoa Part II Ciliophora: Oligohymenophora and Polyhymenophora" Cambridge University Press, 1983 (ref. ID; 2014)
[ref. ID; 3420]
Body conical, irregularly ovoid or posteriorly elongated. In most species somatic ciliature is absent, trichites form a characteristic band, and polysaccharide plates or platelets cover posterior half of body. Predominantly in marine habitats though several common fresh-water forms. (ref. ID; 3420)
Leegaard (1915) showed two forms, slender and rotund, of this ciliate. The posterior extremity is pointed and membranelles of the adoral zone are short. Distinct polygonal cortical platelets cover the body. The macronucleus is round situated in the posterior area. A marine species found in the Atlantic Ocean and the North Sea. (ref. ID; 3420)
Body truncate to conical, 28-53 um in length by 27-44 um in width. Anterior polykinetid zone (APZ) distinctly separated from ventral polykinetid zone (VPZ); APZ comprised of 16 (13-18) polykinetids of equal length surrounding anterior end; VPZ comprised of 9-15 polykinetids lying in a shallow ventral groove that forms an enclosed sunken pit posteriorly. No paroral kinety observed. Girdle (G) of monokinetids with very short cilia, supraequatorial, completely surrounding the cell. Ventral kinety not observed. One spherical macronucleus, 9 (8-15) um in diameter. Large "trichites", usually numerous, inserting above or near G and extending internally posteriorly, often extruding to form beaded strands among cilia of APZ. (ref. ID; 1032)
The body is conical, its length and width are approximately the same. Its posterior extremity is pointed. The macronucleus is oval. Rarely found in the Atlantic Ocean and the North Sea. (ref. ID; 3420)
The body is dorso-ventrally flattened, somewhat elongated and bluntly rounded at the anterior and posterior extremities. The middle area is constricted. This species is characterized by its extreme alveolar or vacuolated appearance. The body seems one mass of globules. A nucleus and contractile vacuole have not been described. A marine species. (ref. ID; 3420)
The peristomial field is inclined because of fixation. Hada (1970) showed with normal morphology although he used a fixative as well before describing the animal. The posterior area is "funnel-like" in shape, covered with polysaccharide plates. In the middle area a different kind of polysaccharide plate is present, which are elongated in shape with round edges at the top. Trichites are present. Busch (1930) suggested that the round granule in the anterior area is probably a food vacuole. It was found in seawater of the Antarctic region. (ref. ID; 3420)
Size ~40x30 um in vivo and ~30x25 um after protargol impregnation. Cell broadly obconical, anteriorly conspicuously slanted leftwards, with peristomial collar. Macronucleus and micronucleus ellipsoidal. Extrusomes acicular and ~15x0.5 um. Girdle kinety usually subequatorial, continuous, composed of ~42 dikinetids. Ventral kinety composed of ~6 dikinetids. Adoral zone composed of average of 16 anterior and 8 ventral membranelles. (ref. ID; 4918)
Descriptions
Body is ovoid. The adoral membranelles are very long and there is a paroral membrane in the peristome. The trichites are long and few in number. The macronucleus is ovoid. It possesses numerous sand grains inside the cell. Found in fine sand in the seashore at Roscoff. (ref. ID; 3420)
Redescription
Size 30-45x25-35 um in vivo, usually ~40x30 um, and 26-34x21-28 um, usually 30x25 um after protargol impregnation. Shape broadly obconical, broadly ellipsoidal or cylindroidal, but usually broadly obconical and circular in cross section; length:width ratio, in contrast, rather constant, namely, 1.1-1.3:1 both in vivo and protargol preparations. Posterior cell end broadly rounded to bluntly pointed; anterior end distinctly slanted leftwards, with thin but distinct peristomial collar decreasing in height from right to left end of anterior membranellar zone, outline thus depends on the side viewed. Macronucleus in left half of cell, ellipsoidal to ovoidal, contains numerous nucleoli 0.5-2 um across; in ~20% of specimens irregular, in a supposed postdivider globular. Micronucleus in shallow indentation of macronucleus, broadly ellipsoidal to lenticular. No contractile vacuole observed. Cortex with polygonal platelets ~3 um across covering cell posterior to girdle kinety, weak in anterior portion of right side, where most cells are damaged in slides. Perilemma usually distended underneath girdle kinety in protargol-impregnated cells, except for region around ventral kinety. Resting extrusomes ("trichites") attached to cortex in shallow bulge above girdle kinety and forming an equatorial funnel, indistinctly clustered, roughly acicular with slightly broadened conical anterior end, ~15x0.5 um in vivo. Ejected extrusomes filiform, i.e. thinner than a cilium in anterior portion, up to 60 um long. Argyrophilic structures (probably fibres) associated with extrusome attachment sites in bulge above girdle kinety, bipartited into a leftwards curved posterior portion and an anterior portion extending obliquely rightwards. Cytoplasm colourless, contains lipid droplets 3-6 um across and food vacuoles 3-7 um across probably with remnants of bacteria and flagellates. Movement very fast and restless, swims straight forward for short distances interrupted by jerks to and fro, while changing direction. Somatic cilia arranged in a girdle and ventral kinety. Girdle kinety ~66% back from anterior cell end, slightly oblique with dorsal portion more posteriorly, continuous, composed of ~42 horizontally orientated dikinetids each bearing two cilia: a rod-shaped, 1.5-2 um long left cilium and a ~0.5 um long right one; the latter recognizable only in scanning electron micrographs. A connecting structure (probably a fibre) and a third granule above each dikinetid impregnate occasionally with protargol. Ventral kinety in posterior sixth of cell, extends in furrow lined by argyrophilic structures (probably fibres) on both sides, composed of ~6 meridionally orientated dikinetids each bearing a rod-shaped, 1.5-2 um long cilium on anterior basal body. Oral apparatus occupies anterior end of cell. Oral funnel eccentric, separated from ventral side by thin cytoplasmic wall, entrance thus between ventral and anterior surface, contains paroral membrane and proximal two thirds of ventral membranellar zone. Anterior membranellar zone almost closed, conspicuously slanted leftwards, surrounds peristomial collar, composed of ~16 um membranelles with cilia up to 25 um long in vivo; membranelles ~1 um apart in protargol preparations, each composed of three rows of basal bodies. System of argyrophilic structures (probably fibres) associated with anterior membranellar zone, composed of (i) a fibre connecting the distal membranelle ends and (ii) a longitudinal fibre between each two membranelles abutting on the connecting fibre with a granule. Ventral membranellar zone set off from anterior zone, almost perpendicular to ventral side, terminates ~35% back from anterior cell end, composed of ~8 membranelles. Ventral membranelles each composed of three rows of basal bodies in distal portion of zone, of only two rows in proximal. Paroral membrane on inner wall of buccal lip, 6-10 um long, composed of probably bare basal bodies as no cilia recognizable in scanning electron micrographs. Pharyngeal fibres insert at proximal end of oral cavity, distinct in vivo and protargol preparations, where they are ~10 um long. (ref. ID; 4918)
Comparison with original description and related species
The description of S. arenicola provides a comparison with S. sauerbreyae and one drawing (Dragesco 1960), but lacks a description and morphometric data, except for cell length (40 um). Thus, a reliable identification will never be possible, and neotypification is required. The cell shape, peristomial collar, extrusomes (~13 um long), membranellar zone (17 anterior and ~8 ventral membranelles), the lack of a contractile vacuole, and the occurrence in marine sediment, however, match my specimens, which thus can serve as neotype. There are some marine Strombidium species with a similar size, nuclear apparatus, and apical protrusion: S. glaciale Petz, Song and Wilbert, 1995 (girdle kinety discontinuous, 9-13 ventral dikinetids, membranellar zone distinctly open): S. purpureum Kahl, 1932 (with purple bacteria); S. sulcatum Clarapede and Lachmann, 1859 as redescribed by Faure-Fremeit and Gainer (1970; membranellar zone not slanted and with 12 anterior membranelles, extrusomes 7-8 um long in TEM); and S. vestitum (Leegaard, 1915) Kahl, 1932, as redescribed by Agatha and Riedel-Lorje (1997; 8-18 ventral dikinetids, girdle kinety pre-equatorial and with ~57 dikinetids). The descriptions of S. rhynchum and S. tressum are based on protargol-impregnated specimens; live observations are lacking (Lynn et al. 1988). Strombidium rhynchum Lynn, Montagnes and Small, 1988 and S. arenicola differ in cell shape (length:width ratio ~2:1, in line drawing even 2.4:1 without distended perilemma vs. 1.1-1.3:1). Although considering that different preparation methods (QPS in Montagnes and Lynn 1987 vs. protocol A in Foissner 1991) influence the cell shape, the discrepancy between S. rhynchum and S, arenicola is rather large, indicating that they are probably no conspecific; a reinvestigation of the former species is, however, required for corroboration. Strombidium tressum has extremely long cilia of the anterior membranelles, whose length is unfortunately not mentioned and also not recognizable in the micrographs provided by Lynn et al. (1988). Ejected extrusome form long (20 um) strands among the "torch-like" spiralled anterior membranelles and thus might have been confused with cilia. Furthermore, the authors did not discuss the similarity between their new species and S. arenicola Dragesco, 1960. Accordingly, the "torch-like" arrangement of the anterior membranelles in the preparations is the sole distinguishing feature, which hardly justifies separation at species level. (ref. ID; 4918)
Ontogenesis: Morphostatic specimens do not show a neoformation organelle. Cell division of Strombidium arenicola can be roughly divided into three stages: early, middle, and late dividers. (ref. ID; 4918)
Early dividers: Stomatogenesis commences with the apokinetal development of a cuneate, longitudinally orientated anarchic field of basal bodies in a shallow depression underneath the girdle kinety and to the left of the ventral kinety. While the oral primordium elongates posteriorly into a subsurface tube, membranelles differentiate from anterior to posterior. When the final number of membranelles has been formed, the oral primordium curves counter-clockwise and orientates inversely to the parental oral structures. Simultaneously, the membranelles become ciliated and the paroral membranelle originates at the bottom of the tube roofed by the ventral membranelles. The somatic kinetids proliferate intrakinetally, and the girdle kinety splits underneath the buccal vertex. A replication band appears in the macronucleus and proceeds posteriorly when the first membranelles have been generated. The micronucleus swells from 1-2 um to 4 um in diameter and divides; the filial products migrate into the anterior and posterior cell halves. Cell length increases by ~1/3 during this stage. (ref. ID; 4918)
Middle dividers: The opisthe's zone of anterior membranelles moves anteriorly to the surface between the circumferential stripe of argyrophilic fibres and the girdle kinety. By the evagination of the opisthe's oral apparatus on the proter's left side, the posterior cell portion, including cortical platelet layer and left end of girdle kinety, is shifted posteriorly, while the ventral kinety does not change position. Next, the girdle kinety splits a second time, i.e. dorsally; the anterior portion extends horizontally across proter's right cell half, while the posterior portion accompanies the lower margin of the emergence slit. The ventral kinety divides, too; it anterior product probably becomes the proter's ventral kinety, the posterior that of the opisthe. Subsequently, the new oral apparatus rights till it is almost parallel to the ventral surface. The macronucleus lengthens, achieving an elongate ellipsoidal shape, while its nucleoli decrease in number and size. The specimens become large by ~25% due to the evagination of the opisthe's oral apparatus. (ref. ID; 4918)
Late dividers: The division furrow becomes recognizable while the opisthe's posterior cell portion forms a distinct bulge on the proter's posterior right side. Then, the opisthe's anterior cell portion commences to enlarge by growing out and the peristomial collar and buccal lip are shaped. The closely spaced kinetids of both widely open girdle kineties will probably diverge to form continuous ciliary rows just before separation. The division products of the ventral kinety diverge by enlargement of the posterior cell portion and migrate slightly to their final positions. The macronucleus becomes bilobate with the nucleoli aggregated in the lobes; finally, it divides into two broadly ellipsoidal nodules each accompanied by a micronucleus. Late dividers enlarge only slightly; postdividers are thus somewhat smaller than the morphostatic specimens. During ontogenesis, no signs of reorganization are recognizable in the parental oral ciliature. (ref. ID; 4918)
The apical protuberance is conspicuous, like a elastic finger in form, usually 9 um in length. The AZM is wide and pinnate in its anterior half. A dense band of trichites is present at the dorsal side, extending from left to end at the right side near the posterior area. One or two contractile vacuoles and an elliptical macronucleus is present. It feeds on diatoms. A marine species. (ref. ID; 3420)
The anterior area is oval and the posterior area is slightly shortened and rounded. The anterior extremity is not covered with polysaccharide plates. The anterior part of the plates is considerably thickened and elongate, rounded at their top. The posterior plates are hexagonal in shape. Near the peristomial field there is a bundle of membranelles in pairs which are probably damaged by the fixative employed. Macronucleus is large and ovoid. This species was frequently found in the Atlantic Ocean. (ref. ID; 3420)
Comments
Busch (1921) placed this species in the new genus Buehringa. But the presence of two different polysaccharide plates are not considered sufficient reason to erect the new genus. (ref. ID; 3420)
The following diagnosis is based on observations of over 30 protargol-stained specimens from Indian Arm, B.C. Fifteen (15-16) anterior and 8 (7-9) ventral polykinetids. Ventral kinety composed of 28 (27-29) dikinetids beginning just posterior to girdle and extending to cell posterior. (ref. ID; 7287)
Descriptions
Cell shape conical with rounded anterior, 41 um (28-52) long and 45 um (37-47) wide. Oral cavity almost enclosed by ring of anterior polykinetids. Anterior polykinetid zone continuous with ventral polykinetid zone. Anterior polykinetids all of equal length surrounding anterior end. Ventral polykinetid zone, comprised of polykinetids becoming smaller posteriorly, lies in ventral groove forming semi-enclosed sunken pit. Ciliated paroral kinety extending down right side of oral groove. Equatorial girdle, believed to be comprised of ciliated monokinetids, completely surrounding cell. Each dikinetid of ventral kinety possesses one short cilium. One sub-spherical macronucleus positioned near center of cell. Trichites, faintly stained, inserting anterior to girdle and extending internally toward cell posterior.
Strombidium basimorphum was represented by two morphs which were observed at different sites. The two forms differed in length only by an average of 15 um, whereas the ranges of width, number of anterior and ventral polykinetids, and the orientation and number of kinetosomes associated with the ventral kinety, were consistent between populations. There is evidence of similar size variations in response to food limitation (Snyder, R.A., Ohman, M.D. & Small, E.B. 1991. Allometric morphological changes in ciliates. J. Protozool., 38:3a). (ref. ID; 7287)
Comparison to related species
The general form of Indian Arm specimens are similar to those of S. acutum Leegaard, 1915, S. conicoides (Leegaard, 1915) Kahl, 1932, S. compressum (Leegaard, 1915) Kahl, 1932, S. crassulum (Leegaard, 1915) Kahl, 1932, S. taylori n. sp. and S. testaceum Anigstein, 1913. Due to poor detail in the description of S. conicoides, comparisons of somatic and oral ciliature are not possible. However, S. conicoides possesses well-defined polysaccharide plates on the posterior half of the cell that appear lorica-like after fixation (Flemming's fixative). Although protargol does not stain polysaccharide plates well, we believe Bouin's fixed material might reveal this lorica-like structure and would indicate distortion due to shrinkage. No such shrinkage was evident in our specimens. Strombidium compressum is much smaller than S. basimorphum (17-28 um vs. 28-52 um long) and possesses a short, inconspicuous ventral kinety compared to the prominent ventral kinety observed for our species. Strombidium testaceum and S. taylori (70-84 um and 21-37 um long, respectively) both share similar dimensions and anterior polykinetid number to S. basimorphum, however, their respective ranges for ventral polykinetids (16 and 12-15) do not overlap with those of S. basimorphum (7-9). Also, both S. testaceum and S. taylori have elongate macronuclei while S. basimorphum has a sub-spherical nucleus. Strombidium acutum and S. crassulum are similar to S. basimorphum in length, width and anterior polykinetid number. However, the orientation of the ventral cavity and ventral polykinetid zone and the number of ventral polykinetids of S. crassulum differ from those of S. basimorphum. Strombidium acutum not only has more ventral polykinetids than S. basimorphum (10-22 vs. 8) but also lacks a ventral kinety. (ref. ID; 7287)
Etymology
The species designation of basimorphum (basis, G.-foundation; morphe, G.-shape) derives from its "typical" strombidiid shape. (ref. ID; 7287)
Type locality
Shallow marine waters (top 5 m; salinity of 22-25 o/oo) of Indian Arm, B.C., Canada (latitude 49 degrees 22'N, longitude 122 degrees 55'W), taken February 1991. (ref. ID; 7287)
Deposition of type material
A holotype of Strombidium basimorphum has been marked on a slide of protargol-impregnated cells (from this field study) and has been deposited in the Ciliate Type Specimen Slide Collection, USNM #43110, U.S. Natural History Museum, Smithsonina Institution, Washington, DC, USA. (ref. ID; 7287)
The body is irregularly ovoid, bluntly rounded anteriorly and narrowed in the posterior area. The peristomial field is large. Apical membranelles are thin and round at the end, like a "rudder and helm". The end of membranelles are occasionally frayed. Polysaccharide plates or polygonal platelets were present but inconspicuous, from the posterior extremity to the area below the peristome. The macronucleus is oval, situated under the peristomial area. A marine species. (ref. ID; 3420)
Brackish and salt water. (ref. ID; 1618)
The body shape is irregularly ovoid and the basal region, hemispheric. The peristomial area, most developed than in Strombidium sulcatum and less deep than in Strombidium lagenula, extends towards the middle of the ventral side. Polysaccharide plates and a transverse cleft are distinct. Trichites make a funnel-like arrangement. There are two long dorsal membranelles, 35-40 um, independent of the adoral zone, the ends of which can stick onto a substratum. The ovoid macronucleus lies in the body in the posterior region above the bundle of trichites. The cytoplasm is hyaline, clear, enclosing various digestive vacuoles, and contains some granulations which are refractile. Marine species. (ref. ID; 3420)
Body 31-61 um in length by 30-55 um in width, squat in shape with large, deep and wide oral groove and thin peristomial collar. Anterior polykinetid zone (APZ) and ventral polykinetid zone (VPZ) clearly separated; APZ composed of 15 (14-16) polykinetids surrounding anterior end; VPZ composed if 14-23 polykinetids lying in a deep ventral groove that forms a sunken pit. One ciliated paroral kinety composed of monokinetids on right side of oral groove. Subequatorial girdle (G) of monokinetids with short stubby cilia, completely surrounding cell. Ventral kinety, posterior and right of the oral region, extending from the girdle to the posterior pole and composed of 5-10 ciliated kinetosomes, often poorly resolved because of placement in ventral longitudinal groove. On macronucleus, variable in shape, but characteristically elongate and shaped like a question mark, sometimes with bulbose terminal portions. "Trichites" present, inserting above G and extending internally posteriorly, possibly being grouped in bundles. (ref. ID; 1032)
The shape of the posterior area is wide and conical. A transparent peristomial collar is characteristically projected. Membranelles of the adoral zone are thick and deployed over a wide area. Polysaccharide plates show wide stripes, among which finer stripes are observed. The macronucleus was not seen. A species rarely encountered in the Atlantic Ocean and the North Sea. (ref. ID; 3420)
Comments
Strombidium capitatum was first described from the north Atlantic Ocean and North Sea. The original description of S. capitatum did not indicate the placement and kinetid structure of the paroral kinety, the shape of the macronucleus, or the presence of trichites. We have amended the original description with detailed information about these structures. The protargol impregnations suggest the presence of a second paroral, lying adjacent to the VPZ. We have been unable to confirm its presence by SEM and have assumed it must represent a skeletal structure. In addition, SEM revealed that a ventral cleft is present, in which the VK lines. This is likely to be the reason Leegaard did not note the VK. (ref. ID; 1032)
This species greatly resembles Strombidium lagenula Faure-Fremiet, 1924. The fine stripes at the posterior area appear to be trichites. (ref. ID; 3420)
The body is transparent, wider in anterior area and truncated at its extremity. The posterior area is narrowed and ends in a distinct tail. A contractile vacuole is situated one-third of the way down the body form the posterior end. A freshwater species. (ref. ID; 3420)
Body conical shaped, 44-61 um in length by 22-35 um in width. Anterior polykinetid zone comprised of 16 (13-18) polykinetids of equal length, surrounding anterior end and not distinctly separated from the ventral polykinetid zone, which is comprised of 10-16 polykinetids, lying in a shallow ventral groove that forms a sunken pit posteriorly. One paroral kinety of ciliated monokinetids on right side of oral groove. Girdle (G), a supraequatorial ring of monokinetids with short stubby cilia, completely surrounding cell. Ventral kinety of 7-15 ciliated monokinetids, restricted to posterior end, to the right of the oral groove. Macronucleus, highly variable in number (1-5), usually three to four, elongate spheroids, approximately equal in size. "Trichites" insert above G and extend part way down cell. (ref. ID; 1032)
Type locality
Salt Pond, in Eastham, MA, USA (41 degrees 50'N, 69 degrees 58'W). (ref. ID; 1032)
Body ovoid or pyriform. Approximately 20 membranelles visible inside the buccal cavity. (ref. ID; 2316)
Body is slender, obovoid and dorso-ventrally flattened, but the frontal area is thick with about 20 membranelles of the adoral zone. In the AZM 3 or 4 membranelles are pointed. The peristomial area is considerable extending to at least one-third of the body from the posterior end. Thrichites are characteristically arranged and make one transverse band from right side to left side in the dorsal area. Macronucleus is an ellipsoid with an associated micronucleus. This animal rotates while advancing and sometimes fixes itself onto the substratum for a considerable time. It feeds on diatoms and is distributed in marine sand and in brackish water. (ref. ID; 3420)
This species has a wide and rounded form, dorso-ventrally flattened. The posterior area is distorted. Membranelles of the adoral zone are rather small, only 1/4-1/3 of the body length. Among them there are four characteristic membranelles which are long and pinnate. A band of trichites, which is sometimes inconspicuous, run from the right to the left lateral side. The macronucleus is reniform. This species is probably a commensal form on Clavelina and it usually moves by creeping. It feeds on small flagellates. A marine species. (ref. ID; 3420)
This species is very similar to Strombidium crassulum but differs in the shape of the membranelles of the adoral zone and the body size. The polysaccharide plate is striped but specimens without stripes were also described. The macronucleus is oval. A rare species from the North Sea. (ref. ID; 3420)
Comments
Laboea emergens Leegaard, 1915 has been synonymised because of the similarity in shape of the body, AZM and polysaccharide plates. Busch's (1921) fixed specimens, S. emergens var. constanziense, has a ventral cleft in the posterior area. (ref. ID; 3420)
This animal is easily differentiated from the polysaccharide plate which displays a conical shape. A marine species. (ref. ID; 3420)
Comments
Leegaard (1915) established the new genus Woodania with this species. However, Kahl (1932) inferred from the diagram that the cell had shrunk due to fixation. As a result, he transferred it to the genus Strombidium. Kahl (1932) mentioned also that this animal resembles S. sulcatum. (ref. ID; 3420)
Body conical, 71-106 um in length by 36-61 um in width, with a shallow oral groove and anterior protruberance (when alive). Anterior polykinetid zone (APZ) and ventral polykinetid zone (VPZ) showing a distinct separation with the latter beginning slightly inside the former; APZ comprised of 16 polykinetids of equal length, surrounding anterior end; VPZ comprised of 10-17 polykinetids lying in a shallow ventral groove that forms a sunken pit posteriorly. One paroral kinety of ciliated monokinetids on right side within oral groove. Girdle (G), a supraequatorial ring of monokinetids, with short stubby cilia, completely surrounding cell. Ventral kinety of 10-25 ciliated monokinetids, restricted to posterior end and to the right of the oral groove. Macronucleus, single (range 1 or 2), highly variable in shape but usually spheroid. "Trichites" inserting in hexagonally packed arrays anterior to G and extending, internally, part was down cell. (ref. ID; 1032)
The yellow coloured body is relatively wide at the anterior and narrowed posteriorly. The band of trichites makes a funnel-like shape. Cortical plates possess longitudinal lines. There is a cirral row near the posterior extremity. The macronucleus is ovoid. A planktonic marine species. (ref. ID; 3420)
In vivo 50-72x33-58 um. Body obconical. Posterior portion with faint stripes parallel to main body axis. Oral cavity extends to girdle kinety. Macronucleus broadly ellipsoidal, contains numerous nucleoli; micronucleus adjacent to macronucleus. Contractile vacuole not recognized. Cytoplasm brownish. Specimens sometimes with small spherical enclosures (endosymbionts?) and clay particles. These particles, found in the posterior portion of living and fixed specimens, were presumably ingested to feed on the attached microorganisms like bacteria, small pennate diatoms and cyanophyceae (Fenchel 1968). Feeds on coccal organisms from various taxa (1-3 um in diameter), centric diatoms (6 um in diameter), pennate diatoms (36x8 um). Swims slowly in spirals with membranelles perpendicular, interrupted by dapping. When it meets an obstacle, it moves backwards. Environmental stress and ageing evoke a distension of the cell surface and thus an enlargement of the cells. Posterior to the girdle kinety, strips and polygonal platelets become visible, which are arranged in rows. The specimens look like S. acuminatum, with the posterior end of the cell curved slightly to the left. Sometimes in individuals of that morphotype, the intermembranellar spaces are covered with bacteria. Extrusomes rod-shaped, 13-20 um long, not grouped, insert in oblique rows in a 4-6 um wide zone anterior to girdle kinety, converge posteriorly. Extrusome girdle interrupted ventrally. Girdle kinety composed of monokinetids, in shallow furrow. Ventral kinety does not extend to girdle kinety, consists of dikinetids, only anterior kinetosome each bearing a 1-2 um long cilium. 15-26 large anterior polykinetids continuous with 10-15 smaller ventral polykinetids. Paroral membrane composed of 29-37 kinetosomes. The oral primordium develops apokinetally above the ventral kinety on a level with the girdle kinety. Strombidium conicum possesses no neoformation organelle during interphase. In early dividers, the oral primordium is comma-like developing in a posteriad direction. Upon reaching the aboral third of the cell, it winds towards the dorsal side. In late dividers, the separated macronuclei move into proter and opisthe. (ref. ID; 4906)
Cyst: The cysts were characterized by a distinctive elliptical shape with a protrusion at one end marking the site of an excystment aperture, which was closed by a hyaline plug or papulla. The cysts with this shape have been termed 'papuliferes'. The papulifere cyst was enveloped by 2 layers; a thin outer layer was set off some distance from thicker inner layer. The inner layer (endocyst) appeared to be a singe rigid wall but the outer layer (ectocyst) was a fragile membranous coat with mucus. Sometimes foreign particles were heavily attached on the outer layer. Using SEM, the outer surface of the inner wall, which appears to be smooth under the light microscope, revealed a network of fine fibers. The outer membranous layer was easily lost during the preparation process for SEM observations. (ref. ID; 1895)
Comments
Lochmann (1908) described Laboea conica from Kiel Bight and the Baltic Sea, where he found the species throughout the year. His drawing show three specimens with a spherical as well as a broadly ellipsoidal macronucleus. The first redescription by Kahl (1932) characterizes the species by the presence of a ventral kinety, a yellowish cytoplasm and a broadly ellipsoidal macronucleus. Using protargol impregnation, Montagnes et al. (1988) described another population with usually a spherical macronucleus and a ventral kinety consisting of monokinetids. The anterior and ventral polykinetid zone show a distinct separation with the latter beginning slightly inside the former. These characteristics differ from those of our population. Although Kahl (1932) doubted that S. acuminatum and S. conicum are conspecific, in vivo observations showed that S. acuminatum is very similar to aged or stressed specimens of S. conicum. Thus, they are considered synonymous. Leegaard (1915) interpreted the reticulated cell surface in big specimens as a phenomenon of ageing, and Busch (1921) considered the distended cell surface to be pathological. (ref. ID; 4906)
Measurements
[Cyst]: The inner wall was 56+/-4.2 um in length and 42+/-2.4 um in width, and the outer layer was 63+/-4.4 um in length and 52+/-3.8 um in width. (ref. ID; 1895)
Size, 50-70 um according to Kahl (1932). Lochmann (1908), Leegaard (1915) and Wulff (1919) reported its size as 20-50 um, 45-75 um and 30-40 um, respectively. These sizes are smaller than that measured by Kahl (1932) probably because of shrinkage by fixative. (ref. ID; 3420)
The body is slender and a conical shape. The posterior extremity shows a button-like protrusion. According to Wulff (1919), trichites are conspicuous and lie close to the somatic pellicle, and polysaccharide plates consist of 10-14 wide longitudinal stripes, among which there are 10-12 fine stripes. The macronucleus is elongated in form, separated into two segments. A marine species found in the Atlantic Ocean and the North Sea. (ref. ID; 3420)
Measurements
Size, 40-50 um and 70 um according to Wulff (1919) and Leegaard (1915), respectively. (ref. ID; 3420)
The body is gradually narrowed from the apical to the posterior area and the posterior extremity pointed. Membranelles of the adoral zone are pinnate. The surface of polysaccharide plates is smooth, covering almost the entire body. Four longitudinal stripes are observed on the ventral side. Trichites make oblique bands, which are located near the peristomial area. The macronucleus was not described. A marine species. (ref. ID; 3420)
Comments
Kahl (1932) suggested that this species was identical to the slender form of Strombilidium acuminatum (Leegaard, 1915) Kahl, 1932. (ref. ID; 3420)
Slender in shape. The posterior area is slightly distorted and its extremity is round. The anterior area is expanded equatorially. Membranelles of the adoral zone are very thick. Polysaccharide plates cover half the body. Trichites are distinct. The protuberance near the posterior extremity indicates a cell just after division. The macronucleus is not described. This species was found in the North Sea and the Atlantic Ocean. (ref. ID; 3420)
The posterior area displays a conical shape and its extremity is sharply pointed. The polysaccharide plate which extends to the middle of the body possesses stripes but no obvious cortical platelets. Membranelles of the adoral zone are very thick and of considerable length. The macronucleus is oval. Rarely described, from the North Sea. (ref. ID; 3420)
The body shows approximately the same length and width. The posterior area takes the form of a cone. Membranelles of the adoral zone are thick, long and short membranelles being raised at the sample place simultaneously. A polysaccharide plate covers half of the body, and possesses no stripe. An oval macronucleus is present in the middle area of the body. A marine species. (ref. ID; 3420)
A cylindrical species and yellow in colour. The size is not mentioned in the original description. According to Kahl's (1932) observation, the body is colorless, 40 um in length. There is a wreath-like furrow in the posterior area. He also suggested that the shape of this animal is variable in several different circumstances. The contractile vacuole is at the right side of the posterior area. It occurs in saprobic in the sea. A poorly described species. (ref. ID; 3420)
The body is deformed by fixation. The peristomial area is projected outward. Membranelles of the adoral zone are very long. There is a oblique furrow at the surface of the posterior body region. The macronucleus is large and round. It was found in the Antarctic current. Hada (1970) also reported the same species in the Antarctic Ocean. (ref. ID; 3420)
The yellow coloured body is obovoid in form with a projected apical collar. A cytoplasmic pad-like protuberance and a transparent thin plate are attached at the left and right side, respectively. Membranelles of adoral zone are pinnate at their anterior extremities. Trichites in a band are present on the dorsal side which runs from the right side, transversely to the left side and extends laterally to the posterior extremity. The macronucleus is round according to Florentin (1901), but Kahl (1932) described it as ellipsoid. It is very active and feeds on diatoms and green algae. A marine species. (ref. ID; 3420)
Laboea conica var. elongata Leegaard, 1915 (ref. ID; 3420)
Descriptions
A slender body with the posterior area tail-like, slightly distorted, probably due to fixation. Polysaccharide plates are stripped and extend to the peristomial area. Many dark coloured granules are present inside the body. A marine species. (ref. ID; 3420)
The body is ovoid and a peristomial collar is present. There are two long thigmotactic cirri in addition to the adoral membranelles. Trichites are numerous on the somatic area. The left lateral side is furnished with fine cirri. The macronucleus is oval and several sand grains are included inside the cell. Polygonal cortical platelets are not seen. A marine species found in the fine sand at Roscoff. (ref. ID; 3420)
Measurements
Size, 75 um (diagram indicates 100 um). (ref. ID; 3420)
The peristomial area is conspicuous. Trichites make an equatorial band in the middle of the body. From the short pointed area at the posterior, long threads extends, sometimes 500 um in length, from which mucous material is produced. Using a such thread the animal attaches to the substratum and rotates or waves freely. This thread can be temporary lost but a new one is easily produced. Another "pin-like" organelle was once observed near the posterior extremity by Kahl (1932). The macronucleus ovoid. A marine species. (ref. ID; 3420)
Measurements
Size, 30-35 um not including the thread-like extensions. (ref. ID; 3420)
The body is globular. The peristomial field is confined to the apical area and adoral membranelles are very fine. Two thigmotactic membranelles are present, apart from the AZM. The band of trichites is conspicuous and the posterior area is covered with polygonal cortical platelets. The large vacant space can be seen at the posterior extremity. The macronucleus is oval, with a small and spherical micronucleus. Several sand grains and diatoms are included inside the cell. This organism is frequently found in marine sand and in brackish pools. (ref. ID; 3420)
Measurements
Size, 50-65 um (diagram indicates 70 um). (ref. ID; 3420)
The body is transparent and a spherical shape. The AZM is confined to the apical area. There are fine cortical speckles on the surface of the body. Contractile vacuoles are large, two in number, and are located posteriorly. A freshwater species. (ref. ID; 3420)
The posterior extremity is slightly pointed. The band of trichites appears like a screwed sash, which starts from the right side of the body, runs obliquely into the dorsal side, and appears again at the posterior extremity of the ventral side. The food consists of brown algae and diatoms. It occurs in brackish water. (ref. ID; 3420)
Measurements
Size, 50 um according to Czapik (1976). (ref. ID; 3420)
The body is small and slender. The posterior area is sharply pointed. The surface of the body displays a number of longitudinal stripes, about 10-13 in total. The macronucleus is ovoid, located near the center. Found in the Antarctic Ocean. (ref. ID; 3420)
Comments
Hada (1970) failed to differentiate between the genus Strombidium and Strobilidium. (ref. ID; 3420)
Anterior cylindrical; slightly tapering toward anterior. Posterior conical. Anterior polykinetid zone slanted toward left side of the cell. No anterior protrusion. Anterior polykinetid (Apk) number: 12-15 (mean 14; SD 0.6). Ventral polykinetid (VPk) number 7-9 (mean 8: SD 0.5). Anterior polykinetid and ventral polykinetid zones almost continuous. Cilia of Apk ~15 um long. Cilia of VPk ~7 um long at anterior and taper to ~4 near cytostome. Paroral kinety present but poorly visible. One slightly ellipsoid macronucleus: diameter 6-12.5 um (mean 8.5 um; SD 1.2). One micronucleus situated near macronucleus; diameter 1.5-2 um (mean 1.9 um; SD 0.2). Girdle of dikinetids (and trichite insertions) almost equatorial. Polygonal cortical platelets present. Ventral kinety extends from posterior pole toward girdle; consists of 7-9 dikinetids with short cilia. Girdle of trichites in posterior radiate from a point near posterior of cell. Trichites ~15 um long, insert ~3-4 um above girdle of cilia. (ref. ID; 490)
Comments
This ciliate lacks an anterior protuberance and is therefore not S. sulcatum Claparede & Lachmann, 1858. It is similar in size and oral polykinetid number to Strombidium compresum (Leegaard, 1915) Kahl, 1932. However, the new species is semi-benthic, has an anterior end which is slanted obliquely in relation to its body axis and has a relatively deep groove containing the ventral polykinetids. On the other hand, S. compresum is planktonic, has an anterior end which is perpendicular to its body axis, and has a relatively shallow groove containing the ventral polykinetids. (ref. ID; 490)
Etymology
The specific epithet inclinatum refers to the slanted anterior end (L. inclin: leaning over). (ref. ID; 490)
A slide, USNM 42120, of protargol stained cells representing hapantotype from Strain CCAP 1680/1 resides in the Ciliate Type Specimen Slide Collection, U.S. Natural History Museum, Smithsonian Institution, Washington D.C., U.S.A. (ref. ID; 490)
Measurements
Length 12.5-30 (mean 23.4; SD 3.8); maximum width 12.5-21 (mean 17; SD 2.7) um. All measurements were from protargol stained material which may decrease cell volume by up to 20%. (ref. ID; 490)
The body is ovoid and cylindrical, narrowed posteriorly. There is a protuberance at the anterior extremity. Trichites in a band are situated at the sides of the body. A marine species. (ref. ID; 3420)
This species was described by Kahl (1932) as Strombidium sp. The body is almost cylindrical but slightly wider at the anterior area. The posterior area is round and bluntly pointed at its extremity. The peristome is not extensive. Fine trichite bands are confined to the lateral areas. The macronucleus is oval. A marine species. (ref. ID; 3420)
Measurements
Size, 70-80 um. (ref. ID; 3420)
Strombidium kryalis Petz, 1994 (ref. ID; 4849 original paper) reported author and year? (ref. ID; 191)
Diagnosis
In vivo about 40-55x40 um. Spherical to slightly ellipsoidal. Single, roundish macronucleus and single, contractile vacuole. Adoral zone of membranelles inconspicuous, surrounds small apical area in closed spiral, consists of 14-17 anterior and 4-6 ventral membranelles which invaginate on anterior surface. 1 equatorial and 1 ventral kinety. 1 equatorial girdle of extrusomes. (ref. ID; 4849)
Descriptions
Outline circular to broadly elliptical, transverse section circular. Posterior half of cell with translucent, polygonal, subpellicular platelets (ca. 1-4 um across), forming firm layer, remaining hemispherical after cells burst. Macronucleus globular to slightly ellipsoidal, in mid-body, contains several nucleoli (ca. 2 um in diameter). Micronucleus globular, in indentation of macronucleus, frequently not impregnated with protargol. Usually, a clear (contractile?) vacuole in left half of cell slightly above mid-body. Cytoplasm contains numerous sequestered chloroplasts, about 1-4 um across, cells thus greenish. On average about 59 pg chlorophyll-a/cell. Chloroplasts functional, a few are retained from cryptophytes, i.e. exhibit orange "phycobiline-epifluorescence", most originate from other microalgae, viz. show red "chlorophyll-epifluorescence" (cp. Stoecker & Silver 1990). Only very rarely, food vacuoles containing small diatoms (about 7 um). Movement rapid, rotating about main body axis. Equatorial girdle (2-3 um wide) around periphery of cell immediately anterior to subpellicular platelet layer; extrusomes insert in this structure. Extrusomes almost parallel to cell surface, not grouped, rod-shaped, 8-13x0.5 um in size, usually not impregnated with protargol. Equatorial and ventral kinety composed of dikinetids, only 1 basal body each with short (ca. 2 um) cilium. Equatorial kinety at edge of platelet layer in or slightly below mid-body, usually continuous on ventral side but sometimes with small gap (preparation artifact?). Ventral kinety short, in posterior third of cell in shallow groove. Ventral adoral membranelles inconspicuous, continuous with anterior adoral membranelles so that adoral zone forms closed spiral around small peristomial field. Anterior membranelles each comprising 3 equally long basal body rows, longest bases about 6 um, cilia 15-20 um. Ventral membranelles not distinctly different from anterior membranelles, invaginate on anterior surface (peristomial field), longest bases about 3 um, others gradually shortened, longest cilia about 12 um; innermost ventral membranelle very likely double-rowed. Anterior and ventral membranelles connected by inconspicuous system of fibres. Oral cavity shallow, funnel-shaped, acentric on anterior surface (defining ventral side), contains in vaginated ventral adoral membranelles and single-rowed paroral membrane, about 6 um long, extending to peristomial field. Oral cavity secluded from ventral surface by cytoplasmic wall. A small bundle of short (ca. 6 um) pharyngeal fibres is sometimes recognizable. (ref. ID; 4849)
Comparison with related species
A "closed" adoral zone of membranelles (circular in top view) and an apically located oral cavity as in S. kryalis are usually characteristics of strobilidiids and tintinnids (Corliss 1979; Kahl 1932; Small & Lynn 1985). In these latter groups the entire "closed" circle is formed by the external adoral membranelles (anterior adoral membranelles). In S. kryalis, in contrast, the anterior adoral membranelles describe an incomplete circle; this circle is completed only by the ventral adoral membranelles. Because there are only few ventral membranelles, the oral cavity of S. kryalis is rather shallow. This is very likely also the reason for its apical location and its seclusion towards the ventral side. In many other Strombidium species, which have a longer ventral adoral zone, the oral cavity is open ventrally and the posteriormost (proximal) ventral membranelles invaginate on the ventral area. Considering this as well as the existence of a subpellicular platelet layer, and extrusome girdle, an equatorial and an ventral kinety, S. kryalis is clearly a strombidiid ciliate. According to the revision of Maeda & Carey (1985) there are no strombidiids with a "closed" adoral zone except S. compressum (Leegaard, 1915) Kahl 1932 and S. globosum Fromental, 1876. Strombidium compressum differs from S. kryalis in its conical shape, the prominent adoral zone, the comparatively large peristomial field, the higher number of ventral membranelles (viz., 6-8) and the monokinetidal composition of the equatorial kinety (Lynn et al. 1988). Strombidium globosum is spherical, similar in size of S. kryalis and also has a very small adoral zone of membranelles. However, it occurs in freshwater and possesses 2 contractile vacuoles (Fromental 1876). Therefore, it is not considered to be conspecific with S. kryalis. None of the few strombidiids hitherto described from Antarctic plankton (Busch 1930; Hada 1970) or sea ice (Corliss & Snyder 1986) matches S. kryalis. These species are different in size and/or shape and have an "open" adoral zone of membranelles. (ref. ID; 4849)
Etymology
The species name is derived from "kryos" (Greek) meaning "ice, icy coldness" and refers to the habitat where S. kryalis was found. (ref. ID; 4849)
Type location
Sea ice floe (about 15x30 m, 72 cm thick) in the Weddell Sea, Antarctica, 69 degrees 12'S, 12 degrees 14'W (core number AN 103119). (ref. ID; 4849)
Type specimens
A hapantotype of S. kryalis as a slide of protagol impregnated cells has been deposited in the collection of microscope slides of the Oberosterreichische Landesmuseum, Linz, Austria. (ref. ID; 4849)
The peristomial lip-like collar is distinctly projected at the apical end and the peristomial field extends to the posterior one-third of the body on the ventral side. The AZM is thickly developed and is constituted of 16-20 membranelles. A band of trichites is situated at the posterior end. Cortical platelets are recognizable. The endoplasm displays a brown colour due to the digestive products. The macronucleus was not observed. This planktonic species is extremely fragile and swims very fast, frequently occurring in the marine environments. (ref. ID; 3420)
The ventral side is flat but the dorsal is heightened. The posterior area is wider and rounded. Membranelles of the adoral zone are numerous, 2 or 3 long membranelles are present. Although the band of trichites is not very conspicuous, it surrounds the body. The macronucleus is oval. Numerous diatoms were observed, packed inside. It moves very rapidly and was found occasionally in marine sand. (ref. ID; 3420)
Measurements
Size, 100-140 um and 110-170 um according to Kahl (1932) and Faure-Fremiet (1950), respectively. (ref. ID; 3420)
Cell conical, with distinctive dorso-ventral flattened posterior end, 70 um (range, 59-83) long, 43 um (32-56) in maximum width (left to right), and 38 um (32-47) in maxium breadth (dorso-ventral). The posterior tapers to a flat, tongue-like end that is curved toward the ventral surface. Anterior polykinetid zone (APZ) and ventral polykinetid zone (VPZ) are continuous. APZ comprised of 15 (13-17) polykinetids, surrounding anterior end. Maximum length of APZ cilia: 23 um (19-26). VPZ comprised of 5 (4-5) polykinetids. One ciliated paroral kinety is on the right side of the oral region. A peristomial collar resides within the APZ and encircles the cytostome. The girdle kinety (GK) is formed of short cilia (~2 um long) spaced 2-3 um apart. The girdle kinety resides in a supra-equatorial, ~5 um below the APZ. The girdle kinety begins just below and to the the right of the VPZ; it runs to the right, across the ventral surface, around the right of the cell, to the dorsal surface, continues around to the left side where it comes to within ~5 um of the kinety origin. A fibre runs above the girdle kinety; on both sides of the gap, produced by the break in the girdle kinety, this fibre bends a right angles and descends to the cell posterior where it arcs across the posterior end of the cell to the cell's left side. The right descending portion of the fibre forms a distinct ridge along the ciliate's right side and creates the tip of a tongue-like projection at the posterior. The left descending portion of the fibre appears to be associated with the ventral kinety. The "ventral" kinety (VK), situated on the posterior end, runs from the cell's left side to ~20% up the cell's right side. The VK is composed of 20-25 dikinetids that possess cilia 2-3 um in length. One elongate bilobed (anterior lobe larger than posterior) macronucleus, 45 (23-58) um long, 19 (15-23) um at maximum width is centrally positioned. In lightly-stained protargol preparations, the macronucleus appears to be formed of 5-8 um vacuoles with dark-staining spheres (~3 um) at the intersections where several vacuoles meet. No micronuclei were observed. Lightly staining "elongate tear-shape" extrusomes (~10 um) insert along the right-posterior side of the descending girdle fibre; these may extrude out of the cell to form fine, moniliform filaments. Trichites insert in a band just above the ciliated girdle and form a conical basket that extends ~10 um into cell. In protargol-stained cells, trichites (or possibly extrusomes) occasionally erupt through the cell above the GK. The tube of the neoformation organelle, which extends to a pouch where the oral anlage forms, opens into the break in the girdle kinety. No contractile vacuole was observed. (ref. ID; 7456)
Notes
Lynn and Small (1998) recognised only four genera in the family Strombidiidae: Strombidium, which has a girdle kinety that forms 1-1.5 turns around the cell; Tontonia, which possesses a tail: Cyrtostrombidium, which has a cytopharyngeal basket: and Laboea, which possesses a girdle kinety that forms a counter-clockwise helix towards the posterior (>2 tunrs), when viewed from the anterior end. A fifth genus, Spirostrombidium, is recognized by Petz et al. (1995); it, like Laboea, has a spiralled girdle kinety. Krainer (1991, 1995) recognised two further genera both of which lack a ventral kinety but differ in the structure of their neoformation organelles: Pelagostrombidium and Limnostrombidium. As the species we describe above has a ventral kinety, lacks a tail and a cytopharyngeal basket, and its girdle only encircles the cell once, this species is in the genus Strombidium. Neither of the two strombidiids that have been observed to form dense blooms (Strombidium sp. (Burkholder et al. 1967) and S. cf. reticulatum, (Dale et al., 1987) are dorso-ventrally flattened, and thus they differ from S. lingulum. There are two free living species of Strombidium, that, like S. lingulum, are dorso-ventrally flattened towards their posterior end: S. cinctum Kahl, 1932 and S. elegans Florentin, 1901. Neither of these species exhibits the characteristic bilobed macronucleus of S. lingulum. Also, the ventral zone of polykinetids in both S. cinctum and S. elegans extends at least one-third of the distance down the cell length, whereas that of S. lingulum is restricted to the anterior end. Strombidium cinctum has an unusual girdle structure, which may place it in the genus Spirostrombidium (Petz et al., 1995); this also makes it dissimilar to S. lingulum. Both S. elegans and S. lingulum have ridges running from near the girdle to the posterior end, but the ridges are on opposite sides: S. lingulum has ridge on the right while S. elegans has it on the left. Furthermore, S. elegans has fewer anterior polykinetids than S. lingulum (~12 vs. 13-17, respectively). These characters distinguish S. lingulum from its nearest morphotype-neighbours and indicate that S. lingulum is a new species. (ref. ID; 7456)
Behaviour
Strombidium lingulum is phototactic: ciliates swam toward a light source in a live sample collected from the bloom. We have no direct indication that this species is mixotrophic; sequestered chloroplasts were not observed, but its phototactic behaviour and red pigmentation are indicators that it may be mixotrophic. There are, however, data that indicate S. lingulum consumes dinoflagellates and other ciliates: a small (~10 um) ciliate was repeatedly observed in S. lingulum at levels of one to eight per ciliate.
Etymology
The specific epithet, lingulum, refers to the tongue-like shape of this species and is derived from the Latin lingula, a tongue. (ref. ID; 7456)
Type location
Hohoae Island, Kyuquot Sound, British Columbia, Canada, 50 degrees 02'03"N, 127 degrees 12'42"W; 0.25 m depth; 14 degrees C; salinity of 31-32 0/00; dissolved oxygen 10.4 mg L-1. (ref. ID; 7456)
Depostion of type material
Two slides of a natural assemblage of protargol-stained Strombidium lingulum reside in the collections of the Natural History Museum, London UK: one slide (registration number 1998:1:22:1) contains a holotype and several paratypes; the second slide (registration number 1998:1:27:1) contains paratypes. (ref. ID; 7456)
The body is oval. The posterior area is tail-like, pointed at its extremity. The circlet of apical membranelles is not closed. The macronucleus is oval. A marine species. (ref. ID; 3420)
Measurements
Size, 80 um. (ref. ID; 3420)
Strombidium lynni Martin & Montagnes, 1993 (ref. ID; 7287 original paper)
Diagnosis
The following diagnosis is based on observations of over 30 protargol-stained specimens from Indian Arm, B.C. Fifteen (14-16) anterior and 18 (15-19) ventral polykinetids. Deep oral cavity with complex pocket and associated ciliated paroral kinety. Ventral kinety beginning ~5 um below girdle, extending posteriorly and spiralling clockwise direction (viewed from anterior) eventually looping back around to ventral surface at posterior end. Multiple (10-18) spheroid macronuclei of varing size. (ref. ID; 7287)
Descriptions
Cell with cylindrical anterior and conical posterior, 47 um (37-56) long by 35 um (28-37) wide. Anterior polykinetid zone distinctly separated from ventral polykinetid zone. Anterior polykinetids all of equal length surrounding anterior end. Ventral polykinetid zone lies in deep ventral groove forming semi-enclosed sunken pit. Two most anterior ventral polykientids distinctly offset from others. Paroral kinety beginning at cytostome runs anteriorly along right side of groove. At groove mid-section, it makes a 90 degrees turn dorsally, following pocket that curves inward to organism's left. Girdle, believed to be comprised of monokinetids with short cilia, completely surrounding cell except for small break on ventral surface. Macronuclei scattered throughout the cell. Numerous trichites inserting anterior to girdle and extending internally to near cell posterior. (ref. ID; 7287)
Comparison with related species
The spiralling somatic ciliature characteristic of the genus Laboea Lohmann, 1908 superficially resembles that of S. lynni. However, it is the girdle in Laboea, not the ventral kinety, that coils around the cell. The cell shape and size of S. lynni are similar to those of S. mirable Penard, 1916, S. testaceum Anigstein, 1913 and S. constrictum (Meunier, 1910) Wulff, 1919. However, each of these lack a sprialling ventral kinety, and possess a single nucleus, lacking the spheroid multiple macronuclei characteristics of S. lynni. Further S. mirable and S. testaceum (60-70 um and 70-84 um long, respectively) are larger than S. lynni (37-56 um long).
Etymology
The species designation of lynni was established to acknowledge the taxonomic and ecological work of D.H. Lynn, Department of Zoology, University of Guelph. (ref. ID; 7287)
Type locality
Shallow marine waters (top 5 m; salinity of 22-25 o/oo of Indian Arm, B.C., Canada (latitude 49 degrees 22'N, longitude 122 degrees 55'W), taken February 1990. (ref. ID; 7287)
Deposition of type material
A holotype of Strombidium lynni has been marked on a slide of protargol-impregnated cells (from this field study) and has been deposited in the Ciliate Type Specimens Slide Collection, USNM #43108, U.S. Natural History Museum, Smithsonian Institution, Washiongton, DC, USA. (ref. ID; 7287)
The body shape is elliptical. The peristome is large and a thick paroral membrane is present. Trichites are more numerous and smaller than those of S. arenicola. The round posterior are is covered with polygonal cortical platelets. The macronucleus is oval, with a small and spherical micronucleus. Several sand grains and diatoms are included inside the cell. This organism is frequently found in marine sand and in brackish pools. (ref. ID; 3420)
The body is pyriform and broadly truncate at the anterior end, the middle of which rises as a small protuberance. The tail is about half as long as the body, structureless, hyaline and sharply pointed. Trichites are abundant in the cortex. The macronucleus is large, spherical and placed in the center of the body. The contractile vacuole lies in the posterior area. This marine species occurs in saprobic places and feeds on Peridinium. (ref. ID; 3420)
Comments
According to Kahl (1932), this animal has three distinct long membranelles among the AZM. The name S. styliferum var. minor was erected by Kahl (1932) in place of S. calkinsi (Kahl, 1932) which was occupied. Kahl (1932) has proposed the new name S. calkinsi after the incorrect identification of a species of Strombidium by Calkins (1901). (ref. ID; 3420)
Measurements
Size, 35 um without the "thorn-like" tail. (ref. ID; 3420)
A yellowish or green coloured species. Similar in shape to S. viride but there is no protuberance at the anterior area. The buccal cavity is deep with a wide lip-like rim on the right side. Small splits are situated at the base of the adoral membranelles. Only 4 bundles of trichites present. Polysaccharide platelets are twice as large as those of S. viride. Macronucleus has an oblique cleft. No tubules encircle the middle area of the body, but an "embryo-like" tube is present in the posterior area. The micronucleus is single and large. Usually zoochlorellae and various Chlorophyceae are seen inside the body. Freshwater species. (ref. ID; 3420)
Comments
Strombidium fallax is yellow coloured. Size, 80 um. The ventral side is slightly and the dorsal side is prominently arched. The middle area of the body equatorially expanded. The peristomial field extends near to the posterior end. Trichites were overlooked by Zacharias (1895) but the transverse furrow at the dorsal side suggests their possible presence. A round shaped macronucleus is situated posteriorly. It feeds on small diatoms and Peridinium. Planktonic animals in freshwater habitats. (ref. ID; 3420)
The ventral side is flat and inclined. The band of trichites arises on the dorsal side and extends to the ventral area. Macronuclei are elongate in shape and separated into two segments. It shows characteristic movement; it waves to and fro and moves convulsively. During locomotion the apical area is not held forward. This species, found in meso-saprobic seawater, is not common. It feeds on flagellates. (ref. ID; 3420)
The body is slender and ogival. The posterior extremity is pointed. In transverse cross section the body is seen to take a lentic form. Two thigmotactic membranelles are recognized in the AZM of C. corsicum. Bands of trichites are present at the lateral and posterior area but are not conspicuous. A small and round macronucleus is situated at the middle of the body. A marine species. (ref. ID; 3420)
Measurements
Size, 30 um according to Entz (1884) but Gourret and Roeser (1888) suggested about 115 um for Clypeolum corsicum although Kahl (1932) was not convinced. (ref. ID; 3420)
The shape varies from pyriform to ovoid or conical, depending on the amount of green inclusion inside the body. The posterior area is narrowed and shortly rounded at it extremity. There is a blunt protuberance at the anterior end. A roughly triangular truncation of the anterior ventral side follows along the peristomial depression. A bundle of trichites forms an internal armature in the shape of a truncated cone. Polygonal cortical platelets cover half the body. Both equatorial and ventral cleft are present. The macronucleus is round or oval. Marine species. This species has a characteristic tidal rhythm concerning encystment and excystment reported by Faure-Fremiet (1948). (ref. ID; 3420)
Resting cyst: 'Frothy plug' covering a circular pylome, cilia still visible at margin of cytoplasm. (ref. ID; 4111)
The body is circular or slightly narrowed posteriorly, and dorso-ventrally flattened. The peristomial area reaches almost to the posterior extremity. At the right side of peristome a lip-like protuberance is present. The macronucleus is very long. It was found originally in the North Sea. (ref. ID; 3420)
An elongated form. The whole body below the AZM is covered with polysaccharide plates. The newly formed daughter cell at the posterior area already has an AZM. Found in the North Sea, but rare. (ref. ID; 3420)
Comments
Although Strombidium oblongum Leegaard, 1915 does not show polysaccharide plates, it has been synonymised with S. ovale because of the resemblance of the body shape and the AZM. (ref. ID; 3420)
Body is slightly deformed but it is hard to designate this animal to a specific taxon. The area of the apical extremity projects as a swelling. In the somatic area there are a number of granules. It was found in the Antarctic Current. (ref. ID; 3420)
The body is oblong, more or less flattened dorso-ventrally, slightly convex at its right side and little concave at its left. The anterior end is broadly truncated in the middle from which rises a process, papillary in form. The posterior end is evenly rounded. The peristomial area extends down obliquely and turns abruptly toward the left into the cytopharynx. The posterior half of the body is furnished with the band of trichites, bordering the periphery to form a distinctly expanded zone which bends from both sides to make an equatorial sash. The macronucleus is elliptical, centrally or subcentrally located. A marine species. (ref. ID; 3420)
The posterior area is slender and narrowed into a cone. In the area where polysaccharide plaques cover the body there are longitudinal stripes and also a long spiral furrow. Macronuclei are present although not clearly described, they may be the dark coloured particles visible, small and oval in shape. Present in the North Sea but rare. Kahl (1932) suggested that the fixative damaged the body, resulting in the formation of the spiral line of polysaccharide plaques. (ref. ID; 3420)
Almost cylindrical in form. The middle area of the body is narrowed where the equatorial cleft is present and the posterior area is wide and rounded. The peristomial field is not very well developed and shows little activity. Trichites are very slender but are distinct and make an equatorial band. Small rod-form purple-bacteria which may be a symbiotic fill the inside of the body. Movement is convulsive but elegant. It occurred in brackish water (salt concentration, 1-2%) and saprobic places where H2S content is high. (ref. ID; 3420)
The body is oval, tapering posteriorly and terminating in an elongate tail. The length of the tail varies from 31 um to 55 um with a width of about 7 um at the base. The peristomial area extends to half the length of the body. The adoral membranelles are not thick. Trichites and polysaccharide plates are not visible. The macronucleus is elongate and situated longitudinally. The micronucleus is single and located in a depression of the macronucleus. There are many food vacuoles in the posterior third of the body, containing fragments of sea weed, bacteria and small diatoms. This animal was found in the intestine of Anthocidaris crassispina. When the intestinal contents are teased into seawater, the ciliate is active and often leaves the substratum to swim into the water. The ciliate tends to go forward in fairly straight lines, but frequently stops, back up and then go forward again, these actions are combined with slow rotation on its longitudinal axis. The backing movement is quick and the tail bends easily when it attaches to another substratum. A marine species. (ref. ID; 3420)
In vivo about 40x30 um. Ellipsoid, prominent protuberance on anterior right end. One discontinuous equatorial and one ventral kinety. Ten to 14 anterior and 8-10 ventral adoral membranelles. One anterior and one equatorial girdle of extrusomes. (ref. ID; 4356)
Descriptions
Outline broadly oval to elliptical, anterior membranelles on prominent protrusion slanted ventrally and leftwards. Slightly flattened dorso-ventrally. Posterior half of cell broadly rounded, with polygonal, subpellicular platelets (ca. 2-4 um across) which are difficult to recognize because of their transparency. Macronucleus ellipsoid, in left half of cell in mid-body, contains small and large nucleoli. Micronucleus lenticular, attached to macronucleus, usually does not impregnate with protargol. Contractile vacuole in upper third of cell, left of ventral membranelles. Cytoplasm with few, colourless, greasy shining globules 2-4 um in size. Feeds on chrysomonads. Movement not particularly rapid, sometimes slightly jumping. Extrusomes arranged in two distinct girdles, indistinctly grouped but apparently not in isolated bundles; not very firmly anchored, easily detached and displaced if cells are arrested for observation. Anterior girdle inserts close beneath anterior membranelles, discontinuous at oral cavity. Equatorial girdle inserts at edge of platelet layer. Extrusomes almost parallel to cell surface, in resting state 12-13x0.5 um, cone-shaped, tapering posteriorly. Several other shapes, probably partially extruded or developing extrusomes with a distended anterior end, have been observed. Triggered extrusomes elongate at posterior end to about 35 um. Equatorial and ventral kinety composed of dikinetids, only one basal body of each with a short cilium (ca. 2 um long). Equatorial kinety at edge of platelet layer, discontinuous on ventral side, slightly spiralling, right end shifted somewhat posterior. Ventral kinety in shallow groove, extends from posterior region almost to equatorial kinety whose right end sometimes appears continuous with it (preparation artifact?); absent (or not stained) in some specimens. Peristomial field conspicuous, surrounded by anterior adoral membranelles each comprising three equally long basal body rows (about 6 um) which bear about 20 um long cilia. Base of left-most anterior membranelle slightly shortened (ca. 5 um). Ventral membranelles continuous with anterior membranelles, invaginated, longest bases about 4 um, comprising three ciliary rows each, longest cilia about 7 um; last two (proximal) ventral membranelles perhaps double-rowed. Oral cavity acentric, with paroral membrane along right border. A complex system of fibers lining the cytopharynx originates from the basal bodies of the paroral membrane and the adoral membranelles. (ref. ID; 4356)
Morphogenesis: Only a few stages were found which resemble those know from other strombidiids (Faure-Fremiet 1953; Kormos & Kormos 1958; Puytorac et al. 1987). They show that the oral primordium originates de novo in a ventral, subsurface position. A paroral membrane forms near the longitudinally aligned, slightly curved adoral anlage. The ventral kinety is resorbed very early (or at lest not impregnated), while the equatorial row still persists. A macronuclear replication band is recognizable. (ref. ID; 4356)
Comments
According to the revision by Maeda & Carey (1985) there is only one marine strombidiid species, Strombidium cinctum Kahl, 1932, which supposedly has two transverse girdles of extrusomes on the dorsal side. However, these are only part of one long extrusome band which extend around the cell in the shape of a number 6 (Kahl 1932). Several other species have helicoidal extrusome bands (Maeda & Carey 1985). There is some resemblance in size, shape, and extrusomes to S. fourneleti, which has, however, two curious thigmotactic membranelles and inhabits marine sand and brackish pools (Dragesco 1960). None of the few freshwater species possesses two extrusome girdles. However, Pelagostrombidium mirabile has two to four supraequatorial bundles, which may resemble a girdle, and a posterior group of extrusomes which is not always present (Krainer 1991; Penard 1916). Penard (1916) mentions that the extrusomes are very easily displaced if cells are slightly squeezed for observation. This indicates that the additional posterior group could be a displaced anterior bundle. Pelagostrombidium mirabile (Krainer 1991; Penard 1916) differs distinctly from S. rehwaldi by the more clearly separated anterior and ventral membranelles, the more posteriorly located extrusome girdle, the exit canal of the neoformation organelle and the embryonic body both of which could not be observed in S. rehwaldi. (ref. ID; 4356)
Etymology
This new species is dedicated to Dipl.-Ing. H. Rehwald (Wasserwirtschaftsamt Munchen), the co-ordinator of the Amper-Sonderprogramm, in appreciation of his continuous support. (ref. ID; 4356)
Type location
River Amper about 2 km downriver from the sewage plant at Geiselbullach, east of Furstenfeldbruck, Germany, 48 degrees 13'N, 11 degrees 21'E. (ref. ID; 4356)
Type specimens
A holotype and a paratype of S. rehwaldi as two slides of protargol impregnated cells have been deposited in the collection of microscope slides of the Oberosterreichisches Landesmuseum in Linz, Austria. Accession numbers: 5, 6/1991. (ref. ID; 4356)
The shape is conical and the lateral view where distinct polysaccharide plaques form a covering show straight lines. This species was infrequently recorded from the North Sea. (ref. ID; 3420)
Long, fusiform body tapering to a slightly curved point posteriorly; 120 (90-134) um long, 30 (27-42) um wide (n=10). The paratene forms a "collar" sloping right to left, enclosing the lower portion of the oral membranelles. Small, irregular, oval macronucleus, 20x10 um. A single micronucleus was observed in some specimens, 1.5 um in diameter, closely associated with the posterior portion of the macronucleus. The ventral kinety extends from the posterior tip anteriorly for approximately one-third of the cell length, then curves to the left. Thirty-two membranelles surround the anterior end and extend down into the oral cavity. (ref. ID; 7603)
Remarks
The taxonomy of this group has been in disarray, as noted in a recent view by Maeda and Carey (1985). This specimens clearly fits the existing criteria for the genus, but does not appear to be similar to any described species. Its "collar", forming a smooth "wrinkle" across the anterior portion of the body, is particularly unique, a feature serving as the basis for the name of the organism. Long pennate diatoms (80-100 um) seem to be the chief food of this species. Other Strombidium species were also present in samples, often as dominant species. The whole group warrants further study: oligotrichs, being soft-bodies, do not fix well. (ref. ID; 7603)
Type locality
Specimens of Strombidium rhyticollare n. sp. were found in sea-ice slush material, sea-ice pore water, and at depths of five meters from open-water column collections from the Weddell Sea, Antarctica. (ref. ID; 7603)
The yellow coloured body is elliptical and dorso-ventrally flattened. The anterior extremity projects bluntly and the posterior end is sharpened. Membranelles of the adoral zone are well developed, their length may reach one-third to a quarter of the body length. The band of trichites shows a characteristic arrangement, which run from the posterior end to both sides. One band turns around from the right side to the left in the middle area of the ventral side. Another extends to the dorsal side and two bands are appear to cross optically at the center of the body. There are small protrusions at the point at which the trichites turn. Small polysaccharide plaques are present, each of which is about 4 um in width. The macronucleus is globular. This species moves in circles rapidly, but sometimes attaches to the substratum with two long thigmotactic membranelles adjacent to the AZM, according to Faure-Fremiet (1950). It feeds on diatoms. Present in marine habitats, especially in sands. (ref. ID; 3420)
Comments
Kahl (1932) and Faure-Fremeit (1950) mentioned animals with different arrangements of the band of trichites. Faure-Fremiet (1950) also described two smaller species, one was very flat and posteriorly sharpened in form, size 50 um and the other was dorso-ventrally flattened and more elliptical, size 75 um. (ref. ID; 3420)
Measurements
Size, 100 um and 80-100 um according to Kahl (1932) and Faure-Fremiet (1950), respectively. (ref. ID; 3420)
This species has a slender ellipsoid form, and the frontal area is flat. The peristome is located longitudinally with a thick lip-like protrusion at its right side. Four long membranelles of the AZM are not pinnate at their end. Trichites are present at the left lateral side. The pad-like organ at the right side of the body possesses creases and a cytoproct-like cleft. The macronucleus is oval and in two sections. Found in meso-saprobic seawater, but not a dominant species. (ref. ID; 3420)
Cell conical, 100 um (range, 80-138) long and 20 um (13-31) wide. The posterior tapers to a fine point that is curved in fixed material. Little separation between the anterior polykinetid zone (APZ) and ventral polykinetid zone (VPZ). APZ comprised of 20 (18-26) polykinetids, surrounding anterior end. VPZ comprised of 7 (6-11) polykinetids which lie in a shallow oral groove. One ciliated paroral kinety on right side of oral groove. The girdle begins just below the oral groove, spirals dextrally (when viewed from anterior) to the dorsal surface, continues around to the ventral surface where it angles posteriorly, spirals to the dorsal surface and extends to ~10 um from posterior. The "ventral" kinety (VK), situated on the dorsal surface, begins 10-20 um from the posterior, runs adjacent to, and on the right (ciliate's right) of, the descending girdle for 5-10 um and then continues to the cell's posterior. VK composed of 8-10 kinetosomes (likely dikinetids). Descending portion of the girdle and VK possess 2-3 um long cilia spaced 2-3 um apart. One ovoid macronucleus, 17 (13-24) um long, 6 (4-8) um wide, positioned one third of the cell length from the anterior end. Lightly staining "trichites" insert along the right side (ciliate's right) of the girdle and extend ~10 um into cell. Dark staining (tear shaped) extrusomes lie along the right side and below girdle. Extruded, moniliform, extrusomes often intertwined with cilia of APZ, extend 10-15 um. (ref. ID; 7330)
Discussion of related species
There are four strombidiid species which, like strain BJLSC, are elongate, conical and ~100 um long: Strombidium cornucopiae (Wailes, 1929) Kahl, 1932; S. pulchrum (Leegaard, 1915) Kahl, 1932; S. rhyticollare Corliss and Snyder, 1986; and Laboea acuminatum Leegaard, 1915 (that Kahl (1932) transferred to the genus Strombidium).
Strombidium cornucopiae is larger than the typical range of strain BJLSC (97-200 vs. 80-138 um) and has prominent striations, which strain BJLSC lacks. Strombidium pulchrum has a spiraling ventral kinety like strain BJLSC, but it is slightly larger (167 um long) and has a more extensive zone of oral polykinetids than strain BJLSC. Strombidium rhyticollare is similar in size to strain BJLSC but differs from it in three ways: S. rhyticollare has 32 oral polykinetids while strain BJLSC has 27 (20+7); S. rhyticollare has a groove around its anterior end (posterior to the oral ciliature) which strain BJLSC lacks; and S. rhyticollare has a ventral kinety that runs one-third the cell length (from the posterior) but does not parallel a descending girdle kinety, as does that of strain BJLSC.
Of the above species, strain BJLSC is most similar to L. acuminatum, although L. acuminatum is smaller (60-88 um). Leegaard illustrated three views of Laboea acuminatum, two of which (Fig.12a, b of (Leegaard 1915)) are similar to strain BJLSC, while the third (Fig.12c of (Leegaard 1915)) is not. Figures 12a and b depict a ciliate with indications of a girdle similar that of strain BJLSC and a similar number of oral polykinetids. Further, the nuclear shape and position, of Leegaard's illustrations of L. acuminatum, are similar to that of strain BJLSC. Any of the older descriptions by Wailes (1929) or Leegaard (1915) could be used to describe stran BJLSC, but the description of L. acuminatum appears most fitting. Further, Wailes (1929) observed L. acuminatum in British Columbia waters.
Kahl (1932), however, transferred a number of species of Laboea to the genus Strombidium; a change that has been generally adopted; the genus Laboea is now restricted to species in which the girdle sinistrally spirals around the cell, and there is a truncated ventral kinety (Montagnes et al. 1988). In his revision, Kahl (1932) moved Laboea acuminatum Leegaard, 1915 to Strombidium acuminatum, but this was already a valid species name: Strombidium acuminatum Stein, 1867. Thus, Strombidium acuminatum (Leegaard, 1915) Kahl, 1932 is a junior secondary homonym, and is invalid. We have provided a preplacement name (nomen novum): Strombidium siculum. (ref. ID; 7330)
Phylogenetic remarks
The structure and position of the somatic ciliature of this species is of phylogenetic importance. Most strombidiids have a "girdle" which encircles the cell and has a slight break on the ventral surface. The organization and phylogenetic origin of this structure have been debated; it has been referred to as a paratene, i.e. kinetosomes of adjacent kineties giving the appearance of alinged kinetosomes (Corliss and Snyder 1986; Krainer 1991; Small and Lynn 1985), as a kinety (Petz and Foissner 1992) or left qmbiguously undefined as a "girdle" (Montagnes and Lynn 1991). In S. siculum, the girdle encircles the cell but the left (the ciliate's left) portion of the kinety descends to the posterior of the cell where it parallels the ventral kinety. Further, there is a fiber (probably a kinetodesmal fiber) associated with the girdle. These two observations suggest that the strombidiid girdle originated from a longitudinal kinety. Likely, this kinety paralleled the oral structures which appear to have undergone sinistral torsion (anteriorly viewed) to form the broken-circle of anterior polykinetids. (ref. ID; 7330)
Etymology
The specifc epithet, siculum, refers to the dagger-like shape of this species and is derived from the Latin sicula, a dagger. (ref. ID; 7330)
Time and locality of isolation
July, from Sechelt Inlet, British Columbia, Canada, 1 m depth, temperature of 20 degrees C, and salinity of 20 0/00 (123 degrees 45'W, 49 degrees 40'N). (ref. ID; 7330)
Deposition of type material
A slide, USNM #47752, of protargol-stained cells representing an hapantotype from strain BJLSC resides in the Ciliate Type Specimen Collection, US Natural History Museum, Smithsonian Institution, Washington, D.C. (ref. ID; 7330)
The anterior area of the body expands, as a swelling, and is considerably distorted. The posterior area is narrowed where three spiraled furrows without cirri or cilia are present. Found in the Atlantic Ocean. (ref. ID; 3420)
The conical body is surrounded with polygonal cortical platelets with 3-6 oblique clefts. The apical collar is low, wide and distinctly retractable. The numbers of macronuclei are 36-72 according to Wulff (1919). Widely distributed in the sea. (ref. ID; 3420)
Measurements
Size, 65-100 um and 70-110 um according to Lochmann (1908) and Wulff (1919), respectively. (ref. ID; 3420)
Yellowish green in colour. The frontal area is widened and the posterior part is cone-shaped. There is a small protuberance at the anterior extremity and a long pointed thorn is present at the posterior end. This thorn seems to be used to attach temporarily onto the substratum. Trichites are very dense which form an equatorial band in the middle area. According to Kahl (1932) the posterior region is covered with polygonal cortical platelets which are extremely small and soft, and the ventral cleft is conspicuous just in front of the stylet. The macronucleus is oblong in shape. It feeds on various groups of algae and small Peridinium. A marine species. (ref. ID; 3420)
Measurements
Size, 70-90 um without the cytoplasmic stylet which is 10-25 um in length. (ref. ID; 3420)
Sensu Faure-Fremiet, 1912: Length 40-50 um. Width maximum 25-30 um. Prominent anterior rostrum. Portion of cell anterior to the girdle is cylindrical; ~25 um. Portion of cell posterior to girdle is hemispherical; ~18 um. Cytostome ventral at bottom of short oral cavity. Anterior polykinetid zone perpendicular to cell axis. Open circle of polykinetids; 12 anterior polykinetids; each 6-7 um wide (anterior polykinetids composed of 20 um long cilia); 7-8 ventral polykinetids. Paroral kinety present. Contractile vacuole present. Macronucleus, subequatorial, in center of cell; nearly spherical about 10 um across. Girdle of dikinetids in transverse groove. Polygonal polysaccharide cortical platelets present. Ventral kinety composed of dikinetids, extends as argentophilic stripe from girdle to posterior pole and onto dorsal surface a short distance. Girdle of trichites in posterior; insert anterior to transverse groove. Habitat: euryhaline, marine, intertidal. (ref. ID; 490)
Sensu Lynn et al., 1988: Material described by Lynn et al. (1988) can no longer be referred to as S. culcatum. Lynn et al. based their diagnosis of S. sulcatum on a combination of its similarity to S. sulcatum sensu Faure-Fremiet & Ganier, 1970 and the absence of an anterior protrusion as they inferred from the original illustrations. Strombidium sulcatum sensu Faure-Fremiet, 1912 has an anterior protrusion. Thus, the species described by Lynn et al. is not S. sulcatum. Lynn et al. noted that, with reference to shape, size and polykinetid number; after S. sulcatum, their species was most like Strombidium emergens (Leegaard, 1915) Kahl, 1932. We conclude that the Strombidium described by Lynn et al. is Strombidium emergens. (ref. ID; 490)
Sensu Fenchel & Jonsson, 1988: Fenchel & Jonsson, 1988 referred to a ciliate as S. sulcatum in an autecological study. Their species and S. sulcatum sensu Faure-Fremiet, 1912 are similar in oral polykinetid number and habitat but differ in size and shape. Most importantly, the former isolate has no anterior protuberance. Below, we describe the isolate referred to by Fenchel & Jonsson. (ref. ID; 490)
The form is ovoid and yellowish brown in colour according to Claparede & Lachmann (1858), but the small form just after cell division is short and conical (Kahl 1932). A protuberance is situated on the apical area, where about 12 membranelles of the adoral zone are massively developed. Conspicuous trichites are located in the posterior one-third of the body, which make a funnel-like arrangement. Polygonal cortical platelets are present but no necessarily conspicuous, and this species possesses a ventral cleft and perilemma. A round macronucleus and a single micronucleus is present. The movement is continuous but not extremely fast. It occurs in saprobic areas in the sea and feeds on bacteria and small diatoms. (ref. ID; 3420)
Comments
According to Faure-Fremiet and Ganier (1970) it possesses highly differentiated somatic ciliature which is possibly specialized in a sensorial function. The diagram of Strombidium minutum displays the dorsal side. The shapes and arrangement of the apical protuberance, polygonal cortical platelets and macronucleus are quite similar to S. sulcatum although the size of S. minutum (30-40 um) is slightly smaller, probably because of shrinkage by the fixative employed. S. minutum and S. sulcatum have been synonymised. (ref. ID; 3420)
Measurements
Size, 30-60 um and 40-50 um according to Kahl (1932) and Faure-Fremiet and Ganier (1970), respectively. (ref. ID; 3420)
The body is an elongate, slender goblet shape. The posterior extremity is pointed. The pellicle is abruptly thickened on the dorsal side and extends transversally to the ventral side. Membranelles of the adoral zone are large and thick, 30 in number. The macronucleus, irregular in shape, is probably highly polyploid. A series of chromatin droplets of various sizes (0.5-8 um) but mainly large can be stained with hematoxylin. This species was found in Strongylocentrotus intermedius as an ectocommensal. A marine species. (ref. ID; 3420)
The body is large and ovoid. Well developed trichites form an inverted triangle. The round and conical posterior region contains numerous granules. It was found in the Atlantic Ocean. (ref. ID; 3420)
The following diagnosis is based on obsevations of over 30 protargol-stained specimens from Indian Arm, B.C. U-shaped macronucleus positioned around oral cavity. Equatorial girdle. Fifteen (13-16) anterior and 12 (12-15) ventral polykinetids. Ventral kinety restricted to posterior of cell, composed of 6-8 kinetosomes. (ref. ID; 7287)
Descriptions
Cell with cylindrical anterior end and conical posterior end, 30 um (21-37) long and 27 um (17-39) wide. Deep oral cavity almost enclosed by ring of anterior polykinetids. Anterior polykinetid zone not distinctly separated from ventral polykinetid zone. Anterior polykinetids all of equal length surrounding anterior end. Ventral polykinetid zone, comprised of polykinetids becoming smaller posteriorly, lies in deep ventral groove forming enclosed sunken pit. Presence of paroral kinety not confirmed. Equatorial girdle, believed to be comprised of monokinetids with short cilia, completely surrounding cell. Ventral kinety suspected to be dikinetids, but no distinguishable. Macronucleus positioned with arms of "U" pointing anteriorly. Numerous trichites inserting anterior to girdle and extending internally toward posterior. (ref. ID; 7287)
Comparison to related species
Strombidium taylori is similar in shape to S. acutum Leegaard, 1915, S. compressum (Leegaard, 1915) Kahl, 1932, S. crassulum Leegaard, 1915, and S. constrictum (Meunier, 1910) Wulff, 1919 but S. acutum, S. compressum, and S. crassulum have spherical or ovoid macronuclei, unlike the U-shaped macronucleus present in the Indian Arm population. Further, S. compressum is smaller than S. taylori (17-28 um long vs. 21-37 um) and possesses a conical rather than cylindrically shaped anterior region. Strombidium crassulum is larger than S. taylori (48-70 um vs. 21-37 um long) and has a more exposed oral cavity. Strombidium acutum, although of similar size to S. taylori (28-52 um vs. 21-37 um long, respectively), possesses large, thickened trichites and lacks a ventral kinety. Strombidium constrictum possesses a U-shaped macronucleus positioned around the oral cavity similar to that of S. taylori. However, S. constrictum has a subequatorial girdle (virtually at the posterior end) which constricts the cell posterior into a "button-like" end. (ref. ID; 7287)
Etymology
Strombidium taylori was named to honor F.J.R. Taylor, Department of Oceanography, University of British Columbia. (ref. ID; 7287)
Type locality
Shallow marine waters (top 5 m; salinity of 22-25 0/00 of Indian Arm, B.C., Canada (latitude 49 degrees 22'N, longitude 122 degrees 55'W), taken February 1990. (ref. ID; 7287)
Deposition of type material
A holotype of Strombidium taylori has been marked on a slide of protargol-impregnated cells (from this field study) and has been deposited in the Ciliate Type Specimen Slide Collection, USNM #43109, U.S. Natural History Museum, Smithsonian, Washington, DC, USA. (ref. ID; 7287)
Strombidium testaceum Anigstein, 1913 (ref. ID; 3420) or 1914 (ref. ID; 1621) reported author and year? (ref. ID; 191)
Descriptions
The body is wide in the anterior region and round or obovate posteriorly. The AZM is distinctive and the ends of the membranelles are frayed. There are narrow creases at the base of the membranelles. Inside the peristome a paroral membrane is present. Bundles of trichites make a equatorial band encircling the body. Polygonal cortical platelets are thick covering the posterior half of the body. The oblique row of short bristles runs from the right side to the left side on the dorsal and extends to the ventral area. The macronucleus is elongate in form. It feeds on small ciliates. A marine species. (ref. ID; 3420)
A small size species (20-40 um) with a yellow coloured body probably due to ingested foods. The apical area bears a short distinct protuberance which is the characteristic feature of this species. The posterior area is slim and its extremity is pointed, which exhibits an elastic contractile movement. Trichites are present below the middle of the body. The macronucleus is ovoid and the contractile vacuole is round. A marine species. (ref. ID; 3420)
Comments
The description of Strombidium acuminatum Stein, 1867 is as follows. The body colour is more or less intensive yellow, and it exhibits a blunt projection at the anterior end and a long cone shape at the posterior. The peristomial area is at the center of the anterior body. Although Stein (1867) did not show the diagram of this animal, S. acuminatum has been synonymised with S. tintinnodes. (ref. ID; 3420)
In vivo about 45-50x45-50 um. Body outline triangular. Macronucleus C-shaped, consists of one to three nodules. Girdle kinety continuous, composed of monokinetids. 15-16 anterior and 17-19 ventral polykinetids. Ventral polykinetid zone curves diagonally across ventral side. (ref. ID; 4906)
Descriptions
Dorsoventrally flattened, cross section thus elliptical. Peristomial lip projects above apical end of cell. Macronuclear nodules contain numerous roundish nucleoli. Micronucleus spherical to ellipsoidal, 3-6 um in diameter (n=2), rarely impregnated with protargol. Contractile vacuole not found. Cytoplasm hyaline, sometimes with red inclusions (endosymbionts?). Feeds on pennate (24x6 um) and centric diatoms (14 um in diameter), the ciliate Myrionecta rubra, and dinoflagellates. Swims slowly and swayingly. Extrusomes inserting in a 2-3 um wide zone immediately anterior to girdle kinety, in resting state about 8 um long. Girdle kinety parallels posterior portion of ventral polykinetid zone, i.e. extends obliquely towards right side. Ventral kinety not found. Anterior membranelles cirri-like, radially projecting. Ventral polykinetid zone continuous with anterior polykinetids. Paroral membranelle on right of oral cavity. Strombidium triquetrum possesses no neoformation organelle during interphase. The oral primordium develops posterior of the oral cavity. During morphogenesis, the oral primordium elongates to the dorsal side and finally surrounds the posterior cell portion almost completely. (ref. ID; 4906)
Comments
As concerns shape, size, movement and fragmentation of macronucleus, S. triquetrum is very similar to S. obliquum Kahl, 1932. However, Kahl (1932) clearly shows a discontinuous, slightly spiralling girdle kinety and extrusome girdle. Furthermore, the ventral polykinetids zone is less prominent, being apparently composed of fewer membranelles. Thus, we do not consider our population to be conspecific. The food vacuole contents show that S. triquetrum feeds on Myrionecta rubra. However, it is unclear, whether the reddish inclusions in the cytoplasm are cleptoplasts derived from M. rubra or digestive products. (ref. ID; 4906)
Etymology
The species name is derived from the triangular (triquetrus, lat. =triangular) body shape. (ref. ID; 4906)
Type locality
Plankton of Meldorf Harbour in the Speicherkoog Dithmarschen (station DH, 54 degrees 5'35"N, 8 degrees 57'8"E; west coast of Schleswig-Holstein). (ref. ID; 4906)
Type specimens
Holotype and paratype as 1 slide each of protargol impregnated cells have been deposited in the collection of microscope slides of the Zoologische Museum Hamburg. (ref. ID; 4906)
Body is globose, slightly conical posteriorly. Its surface does not show the furrow-like form of trichites. The circlet of apical membranelles is not completed, but rather forms a spiral arrangement. The peristomial are shows a crescent-shaped cut like the freshwater oligotrich Halteria grandinella. Freshwater species. (ref. ID; 3420)
Comments
Claparede & Lachmann (1858) were unsure if this species belonged in the genus Strombidium. Kahl (1932) transferred it to the genus Strombidinopsis, as a synonym of S. gyrans. Based on the description of Claparede & Lachmann (1858) and a diagram of Roux (1901), this species has been placed in the genus Strombidium. (ref. ID; 3420)
The body shape is ovoid, a peristomial collar present at the anterior end. The posterior extremity is bluntly pointed. A peristomial field extends almost to the posterior end. Membranelles of the adoral zone are thick. A considerable quantity of granules exists inside the body. Marine species.
Comments
This species seems to be similar to Strombidium lagenula, but shows a larger size and possesses no transverse cleft or trichites. (ref. ID; 3420)
The body is a bulky, almost pot-like in shape. There is the projected peristomial collar in the apical area. The AZM is abruptly bent to the left side at its posterior half, which includes three characteristic long membranelles for attaching the body to the substratum. The macronucleus is oval. A contractile vacuole is present posteriorly. This species was found in the Baltic Sea. (ref. ID; 3420)
Comments
This species was described by Stein (1867) without a diagram. Faure-Fremiet (1932) first produced the diagram of this animal which was sent to him from E. Maupas as a personal communication in 1907. (ref. ID; 3420)
The form is turbinate, varying to obovate and broadly elliptical. The peristomial field extends along the ventral side to nearly half the body length. Membranelles of the adoral zone are thick, about half has long as the body. The body is colourless and its surface is smooth without supplementary cilia. There is a slender thread at the posterior extremity for fixing to the substratum, this is contractile and contains a gelatinous substance. The macronucleus is irregularly spherical situated sub-centrally, and the round contractile vacuole is in the anterior area. The movement is extremely rapid and erratic, frequently gyrating for a time around a fixed point, then suddenly darting away. The food consists of diatoms. Found in a pond water with Vaucheria. (ref. ID; 3420)
The following diagnosis is based on observations over 30 protargol-stained specimens from Indian Arm, B.C. Fifteen (14-16) anterior and 6 (5-7) ventral polykinetids. Anterior polykinetid bases all of equal length except for one at left side of entrance to oral cavity that is curved, its internal and external ends pointing posteriorly. Ventral kinety composed of 11 (10-14) dikinetids of which the right kinetosome is ciliated. (ref. ID; 7287)
Descriptions
Cell with conical posterior end and cylindrical anterior end, 27 um (21-33) long and 19 um (14-22) wide. Deep oral cavity extending down to girdle level, almost enclosed by ring of anterior polykinetids. Anterior polykinetid zone distinctly separated from ventral polykinetid zone. Ventral polykinetids all of equal length except for the most posterior, which is ~1/2 length of others. Ventral polykinetids lie in deep ventral groove forming semi-enclosed sunken pit. Ciliated paroral kinety extending full length of oral cavity along its right side. Girdle, believed to be comprised of ciliated monokinetids, completely surrounding cell. One ovoid macronucleus positioned near center of cell. Trichites not seen within cell but insertion points present just anterior to girdle. (ref. ID; 7287)
Comparison to related species
There are a number of long, cone-shaped strombidiids that look superficially like S. ventropinnum: S. tintinnodes Entz, 1884, S. typicum (Lankester, 1874) Butschili, 1889, S. rhynchum Lynn et al., 1888, S. conicum (Lohmann, 1908) Wulff, 1919, S. chlorophilum Montagnes et al., 1988, S. dalum Lynn et al., 1988, and S. tressum Lynn et al., 1988. However, S. tintinnodes, S. typicum and S. rhynchum bear distinct anterior protuberances that S. ventropinnum lacks. The oral and somatic ciliature of S. ventropinnum are similar to S. conicum and S. chlorophilum. The latter two, however, are larger (71-86 um and 44-61 um long, respectively) and possess more ventral polykinetids (10-17 and 10-16). In addition, the macronucleus of S. chlorophilum is comprised of three to four elongate spheroid segments, contrasting to the single macronucleus present in our species. Strombidium conicum also possesses a less distinct and shallower ventral groove. Strombidium dalum and S. tressum can be distinguished from S. ventropinnum by their absence of a deep ventral groove. Strombidium dalum also lacks a ventral kinety and usually possesses a conical macronucleus positioned at the posterior of the cell. In addition, S. ventropinnum lacks the long cilia of the anterior polykinetids and associated extruded trichites present in S. tressum. (ref. ID; 7287)
Etymology
The specific epithet, ventropinnum (ventro, L.-ventral; pinna, L.-feather or plume) derives from the distinct ventral polykinetid zone and associated ventral polykinetids. (ref. ID; 7287)
Type locality
Shallow marine waters (top 5 m; salinity of 22-25 0/00) of Indian Arm, B.C., Canada (latitude 49 degrees 22'N, longitude 122 degrees 55'W), taken February 1991. (ref. ID; 7287)
Deposition of type material
Holotype and paratype specimens of Strombidium ventropinuum have been marked on a slide of protargol-impregnated cells (from this field study) and has been deposited in the Ciliate Type Specimen Slide Collection, USNM #43111 and #43135, respectively, U.S. Natural History Museum, Smithsonian Institution, Washington, D.C., USA. (ref. ID; 7287)
In vivo 20-35x15-30 um. Body shape obovoid to obconical. One continuous girdle kinety in anterior third of cell and one ventral kinety. 13-18 anterior and 5-8 ventral polykinetids. Ventral polykinetid zone distinctly separated from anterior polykinetid zone, which forms an almost closed circle. (ref. ID; 4906)
Descriptions
A very small form. Almost the whole body is covered with polysaccharide plates. The equatorial region is shallowly constricted. The macronuclei are not conspicuous. This species was found in the various areas of the North Sea. (ref. ID; 3420)
Redescription
Longer than wide and slightly flattened, right buccal lip slightly projecting. Posterior cell surface sometimes distended, reticulation not visible in vivo, in Lugol fixed specimens, however, indistinct polygonal platelets were sometimes observed. Macronucleus ellipsoidal, in mid-body, contains numerous nucleoli. Micronucleus spherical to slightly ellipsoidal anterior to or laterally adjacent to macronucleus. Contractile vacuole was not found. Cytoplasm hyaline, often contains numerous food vacuoles. Feeds on centric diatoms (3-4 um in diameter), coccal organisms and flagellates. Extrusomes not grouped, insert in diagonal rows, converge posteriorly. Extrusome girdle parallels girdle kinety, 1-2 um wide. Girdle kinety composed of dikinetids, only left kinetosome each with short stubby cilium. Ventral kinety in shallow groove, composed of dikinetids, not extending to girdle kinety, only anterior kinetosomes ciliated. Ventral polykinetid zone commences distinctly inside of anterior polykinetid zone. Cytostome rather centrally located. Distance between anterior polykinetids 1-2 um. Paroral membrane not found. Strombidium vestitum possesses no neoformation organelle during interphase. The oral primordium originates apokinetally left of the ventral kinety. During morphogenesis, the oral primordium elongated to the dorsal side and finally surrounds the posterior cell portion almost completely. Simultaneously, the macronucleus develops a replication band and elongates. (ref. ID; 4906)
Comments
Based on fixed material, Leegaard (1915) described two very similar species differing only in size, viz. Strombidium vestitum (Leegaard, 1915) Kahl, 1932 and S. delicatissimum (Leegaard, 1915) Busch, 1921; the latter is slightly smaller (22-30x17-25 um vs. 15-20x12-16 um); Busch (1921) even mentioned 9-13 um. Thus, our population is more similar to S. vestitum. However, we found also considerably smaller specimens than these, suggesting that S. delicatissimum is a synonym of S. vestitum. Fixed specimens from the North Atlantic, which resemble S. vestitum, show a reticulation of the posterior cell surface (Busch 1921). Consequently, Busch (1921) considered S. vestitum to be a synonym of S. reticulatum. However, the latter species is twice to three times larger. Therefore, we do not regard it as conspecific. Strombidium vestitum differs from S. sulcatum Claparede & Lachmann, 1858 in a more anteriorly situated girdle, a less prominent protuberance and the absence of a contractile vacuole. It differs from S. tressum Lynn et al., 1988 by the absence of exceptionally long anterior polykinetids. Strombidium inclinatum Montagnes et al., 1990 is distinguished by the almost continuous anterior and ventral polykinetids, the equatorial girdle kinety, and the macronuclear shape. (ref. ID; 4906)
Type specimens
Neotype as 1 slide of protargol impregnated cells has been deposited in the collection of microscope sides of the Zoologische Museum Hamburg. (ref. ID; 4906)
Size, 28-38 um. (ref. ID; 3420)
No diagram is available. Very similar to Strombidium constrictum, but the button-like protuberance at the posterior extremity is not present. The longitudinal stripes, 4 um in width, are clearer in the anterior area than those in the posterior. Each sixth stripe extends to the pointed posterior extremity. (ref. ID; 3420)
Measurements
Size, 40-50 um. (ref. ID; 3420)
Strombidium viride Stein, 1859 (ref. ID; 1219, 1621, 1629) or 1867 (ref. ID; 3420, 4488) or (Stein, 1867) Krainer, 1995 (ref. ID; 4613) reported author and year? (ref. ID; 3954)
See
Limnostrombidium viride (ref. ID; 4613)
Synonym
Strombidium nasutum Smith, 1897 (ref. ID; 3420)
Descriptions
The anterior part of the body is shaped like an inverted cone and there is a blunt, unsharpened protuberance at the anterior extremity. The posterior area is slightly elongated, conical in form. Bundles of trichites make an equatorially encircling band in the middle area of the body and several tubules are encircled with 3-4 folds in the same area. Polygonal cortical platelets of polysaccharide composition are extremely small, around 1-2 um in size. Numerous green coloured cells are described, inside the body. Freshwater species. (ref. ID; 3420)
Comments
Stein (1867) did not show a diagram of this animal, however Penard (1920) and Kahl (1932) redescribed it with a detailed diagram, although Penard's (1920) S. viride has no protuberance in the anterior area. Roux's (1899) S. viride is different from Penard's (1920) and Kahl's (1932) on the shape of the AZM and the body. Kahl (1932) observed three types of S. viride. (1) Large round form (size, about 80 um) with dense colour from zoochlorellae, and green algae inside the body. Abundantly distributed among aquatic plants. (2) Slender form, size about 60 um. The posterior area is gently pointed. Found in deep lakes. A green coloured body with zoochlorellae and their fragments (3) Small and round form, size 40-60 um. Yellow coloured species without zoochlorellae inside the body. Distributed in deep lakes. Strombidium nasutum was found in pond water, size about 50 um in length. (ref. ID; 3420)
Measurements
Size, 60-80 um and 40-80 um according to Penard (1920) and Kahl (1932), respectively. (ref. ID; 3420)
The body is elongate and conical with stripes in the somatic area. The peristomial field is distinct. Ten to twelve trichites are present situated obliquely near the equatorial cleft. Polygonal cortical platelets cover three quarters of the body. About 18 macronuclei are present. A marine species. (ref. ID; 3420)
Comments
Wulff (1919) described this species under the name S. striatum. This was later changed by Kahl (1932) as it was clear that Wulff (1919) had described a new species. (ref. ID; 3420)