Paramecium
Paramecium O.F. Muller, 1773 (ref. ID; 2014)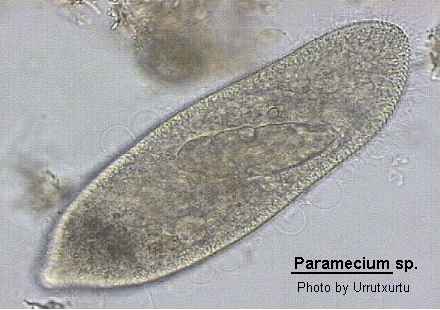
Ciliophora: Hymenostomatida (ref. ID; 7599) Synonym Paramaecium O.F. Muller (ref. ID; 1618) [ref. ID; 2014]
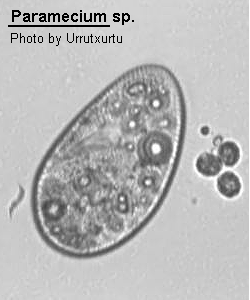
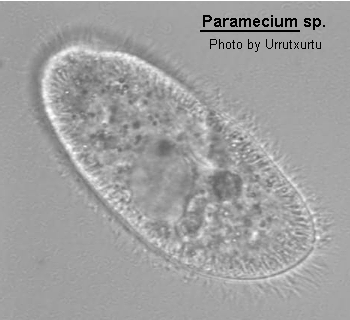
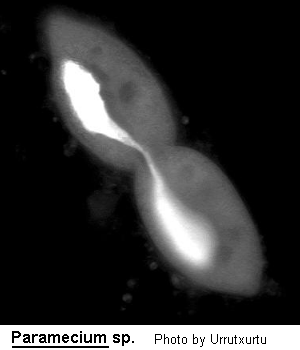
Medium- to large sized ciliate (120-300 um long), ovoid, elongate, foot- or cigar-shaped. The anterior end may be rounded or obliquely truncated and the posterior rounded or conical. The shape is rather asymmetrical due to the presence of an oblique depression, the oral groove, running from the apex of the cell to about the middle of the body on the ventral surface. The oral aperture is located at the posterior end of the oral groove. While the oral groove is prominent and easily seen the aperture is rarely visible without staining the cell with silver. Inside the buccal cavity there is a comparatively small undulating membrane on the right and 2 membranelles called peniculi lying alongside each other. Membranelle M3 comprise four rows of kinetosomes which diverge centrally to form a structure known as a quadrulus. Somatic ciliation is complete and uniform with 1, 2 or 3 hexagonal units. There are both pre- and post-oral sutures and trichocysts present. 1 to many (commonly 2) contractile vacuoles present which may be served by radiating canals. Cytoproct on the ventral surface located sub-orally. Macronucleus ovoid to reniform in shape, usually centrally located with variable number of micronuclei close by. Locomotion characteristically vigorous and spiralling anticlockwise. Wenrich (1928). The species of Paramecium are still in a state of flux. The classical species P. aurelia has recently been subdivided into 14 sibling species by Sonneborn (1975) and Gates, Powelson and Berger (1974) and Gates and Berger (1976) have discussed the morphological separation of these species. Vivier (1974) also reviews the classically named species.
Quote; Colin R. Curds, Michael A. Gates and David McL. Roberts "British and other freshwater ciliated protozoa Part II Ciliophora: Oligohymenophora and Polyhymenophora" Cambridge University Press, 1983 (ref. ID; 2014) [ref. ID; 7435]
The molecular diversity within the D2 domain of the 23S ribosomal RNA molecules of Paramecium. (ref. ID; 7435)
- Paramecium africanum (ref. ID; 4903)
- Paramecium ambiguum (ref. ID; 4903)
- Paramecium arcticum (ref. ID; 4903)
Paramecium aurelia-complex (ref. ID; 4061, 4062, 4063, 4064, 4611, 5910, 7704)
[ref. ID; 4061]
Traditional means of distinguishing different species of the genus Paramecium have been by body and nuclear morphology. The discovery of mating-type characteristics by Sonneborn, however, led to the realization that what was though to be a single species (Paramecium aurelia) was, in fact, at lest 14 genetically isolated sibling species. Sonneborn hesitated to establish separate "named" species for many years because of a lack of objective criteria for establishing mating-type groups. This difficulty was overcome in the early 1970's when isoenzyme gel migration characteristics showed correlations with such groups (which Sonneborn had termed "syngens"); this allowed the establishment of formally named taxonomic species. (ref. ID; 4061) [ref. ID; 4063]
Originally, the taxon "Paramecium aurelia" was used to lump together a number of organism that were morphologically similar but between which gene flow did not always occur. As more information became available, particularly through application of biochemical techniques, classification of this group of organisms achieved finer resolution. At first, the 14 subgroups were called "varieties" on the basis of specificity of the mating types, that is, the specific selectively of the breeding relations. Subsequently the 14 varieties were designated "biological species", or "syngens", but not given species status, on the grounds that mating type identification was difficult to learn and apply to new strains and that it was undesirable to base identification on living material that had to be maintained in the laboratory and could easily be lost or mixed up. When it became possible to freeze and store paramecia in liquid nitrogen efficiently, and the syngens could be identified by simple biochemical procedures (such as starch gel electrophoresis of enzymes), species status was at last given to the 14 syngens, now considered sibling species of an "aurelia complex". (ref. ID; 4063)- Paramecium primaurelia (ref. ID; 4062, 4063, 4611) = Paramecium aurelia subspecies 1 (ref. ID; 3829)
- Paramecium tetraurelia (ref. ID; 4062, 4063, 4611) = Paramecium aurelia syngen 4 (ref. ID; 3737, 3811)
- Paramecium biaurelia (ref. ID; 3987, 4062, 4063, 4611)
- Paramecium decaurelia (ref. ID; 4063, 4611)
- Paramecium dodecaurelia (ref. ID; 4062, 4063, 4611)
- Paramecium octaurelia (ref. ID; 3987, 4062, 4063, 4611)
- Paramecium novaurelia (ref. ID; 4062, 4063, 4611)
- Paramecium pentaurelia (ref. ID; 4062, 4063, 4611)
- Paramecium quadecaurelia (ref. ID; 4063, 4611)
- Paramecium sonneborni (ref. ID; 4061)
- Paramecium septaurelia (ref. ID; 4062, 4063, 4611)
- Paramecium sexaurelia (ref. ID; 3987, 4062, 4063, 4611)
- Paramecium tredecaurelia (ref. ID; 3987, 4062, 4063, 4611)
- Paramecium triaurelia (ref. ID; 4062, 4063, 4611)
- Paramecium undecaurelia (ref. ID; 4062, 4611)
The organism known for nearly 200 years as Paramecium aurelia was only one of several commonly recognized species of the genus. From the point of view of easily determined phenotypic characters (body size and shape, number of micronuclei, number of contractile vacuoles, presence or absence of symbitotic zoochlorellae), it was generally readily distingushiable from such other frequently encountered forms as P. caudaum, P. multimicronucleatum, and P. bursaria. Determination of these "morphological" species was possible by use of simple light microscopy, with almost any textbook as guide (e.g., Kudo 1966). The pre-1975 taxonomic history of Paramecium spp. is well documented in many places: the aging but author-itative accounts by Kalmus (1931) and Wichterman (1953) may be cited as well as the relatively recent chapter by Vivier (1974). Nearly 45 years ago, Sonneborn (1939), appreciating the abiding genetic differences he had discovered among various lines of P. aurelia, proposed labels that could be used of separate such different breeding populations. The term he suggested was "variety", and it was immediately adapted by ciliate geneticists and physiologists everywhere. Some 18 years later, keenly aware that he was dealing with "physiological" and/or "genetic", as opposed to the classical "morphological", species, Sonneborn (1957) introduced the word "syngen" to better describe what he and others had been calling varieties. Although general protozoologists were often reluctant to accept this term (excellent though it was) and some ecologists (e.g., Hairston 1958) chided the Bloomington school for not assigning at that time regular latinized names to these sibling or cryptic species, Sonneborn, with commendable caution, postponed such a significant taxonomic step (i.e., splitting up the long familiar P. aurelia into several separate species all with different names) until he felt certain that more relief than confusion would result from such an action. Then, in 1975 (though see also Sonneborn 1974), convinced that numerous characters (beyond only sexual compatibility ones) were finally available for distinguishing paramecium species and that living "reference specimens" of each were obtainable by anyone from a reliable long-term source, Sonneborn took the overdue step and published his already classic paper (Sonneborn 1975) on the "Paramecium aurelia complex of fourteen sibling species", in which he gave fully distinctive taxonomic names to each. In short, varieties of the old P. aurelia had become syngens that, in turn, now became taxonomically valid separate species. (ref. ID; 7704)- Taxonomic-nomenclatural consequences: Although Sonneborn's decision was laudable and offered neither surprise nor difficulty to co-workers in ciliate genetics, it is still having far-reaching effects on the thousands of other biologists around the world who, on occasion, work with or have to deal with species of Paramecium. The list of such persons includes teachers, all levels of students, textbook writers, review authors, and editors, as well as ecologists, evolutionary biologists, taxonomists, hydrobiologists, cytologists, cell physiologists, and biochemists. Since the essentially irreversible consequences of Sonneborn's action affect many biologists, though some may be quite unaware of this fact, what he actually did warrants further clarification and discussion.
- Names of the new species involved. Currently, there are 14 "sibling species" comprising the Paramecium aurelia complex (shortenable to the "aurelia complex" when it is clear that these paramecia are the topic). These are valid taxonomic species, and their names were both wisely and ingeniously based on the numerical sequence used -with rare exception (see Sonneborn 1975)- in the older designations of variety and syngen. The new names follow; the authorship and date of each is the same, viz., "Sonneborn, 1975": Paramecium primaurelia, P. biaurelia, P. triaurelia, P. tetraurelia, P. pentaurelia, P. sexaurelia, P. septaurelia, P. octaurelia, P. novaurelia, P. decaurelia, P. undecaurelia, P. dodecaurelia, P. tredecaurelia, and P. quadecaurelia. These are the specific names for former syngnes numbers 1-14. Over the years, most intensively studied of these species in genetic research has been P. tetraurelia (i.e., old variety, later syngen 4). (ref. ID; 7704)
- What happened to P. aurelia itself?. A major problem that confronted Sonneborn for some 30 years, as he contemplated separating the several species of "P. aurelia", was what to do with the original single taxonomic name. He was urged by many to retain it: certainly textbook writers and general biologists would have appreciated having that time-revered and so widely used a name preserved (it is found in titles of books, e.g., see Beale, 1954, and Jurand & Selman, 1969, as well as in hundreds of papers). But, in brief, the dilemma he faced was this: If one syngen (e.g., number 1 or number 4) was deliberately chosen to be the one-and only P. aurelia, then how or what could the remaining sibling species be named? The judicious idea of using the sequential-numerical latinized prefixes listed above would have to go, since a "hole" would be caused by removal of either 1 or 4. Furthermore, and this is supported by provisions of the Internationl Code of Zoological Nomenclature (see Corliss 1963) how can anyone known exactly which syngen O.F. Muller was looking at two centuries ago, when he used the specific epithet "aurelia" for this ciliate? So, Sonneborn (1975) declared "P. aurelia" to be an unusable specific name, not attachable with certainty to an organism identical with O.F. Muller's ciliate, thus truly a nomen dubium. In effect, then, P. aurelia, as a valid taxonomic name, has disappeared. Because of overlapping in morphological features, general descriptions of all 14 new species essentially fit the older characterizations of the single P. aurelia. For this reason, we urge that the term "P. aurelia complex" be used in the general literature of the future as the best single replacement of the original, now on-existent, species. Today, specialists can, and should when appropriate, go a step further and state their papers, when they know it to be true, that species P. primaurelia and P. tetraurelia (or whatever) of this complex were the specific organisms investigated in their particular study. (ref. ID; 7704)
- Characteristics used to separate species: Before 1970, syngen (now species) separation depended almost entirely on breeding tests, demonstration of the failure to obtain gene flow between any two groups that would result in viable (and lasting) populations of offspring. Thus, proper identification depended on having standardized living cultures ("tester" stocks) available for the tests and on possessing the requisite technical knowledge to handle the organisms in a precise and controlled manner (Sonneborn 1950, 1970). College teachers and field ecologists were not usually expected to go to such trouble for their routine requirements in identification: "Paramecium aurelia" adequately met their needs. Now, the situation has changed. Presumptive identification of all species can be routinely carried out without recourse to standard living cultures, thankns principally to the availability of techniques for enzyme analyses (Adams and Allen 1975; Allen 1967; Allen and Gibson 1975). Newer morphological information, including recognition of specific endosymbionts, and data on zoogeographical distribution have also contributed characteristics usable by biologists in distinguishing unambiguously at least some of the species comprising the aurelia complex. Finally, purebred stocks (generally kept cryopreserved), serving as standard or reference strain material, are now conveniently available to anyone who may wish to fully match collections from the wild with pedigreed known material. (ref. ID; 7704)
- Isozyme patterns. Under standardizedd conditions, the isozymes of an enzyme for a given organism may present a consistent distribution pattern (a zymogram) if the enzyme is electrophoresed through a starch gel. Since the expression of isozymes is a phenotypic expression of the genotype of an organism, enzymes can be studied geneticists like any other expressed heritable trait. Beginning in the late 1960's and continuing into the '70's, S.L. Allen and other investigated enzymes of paramecia now assigned to the aurelia complex (Admas and Allen 1975; Allen et al. 1971, Allen et al. 1973; Allen and Gibson 1971, 1975; Byrne and Cronkite 1971; Tait 1970, 1978). One result of many of these studies was the demonstration that esterase zymograms could be used to recognize differences among all of the species except two, P. primaurelia and P. pentaurelia. More recently, using the same approach as Allen and her colleagues for 172 bacterized strains of the aurelia complex, it has been shown to be possible to distinguish all species (including P. primaurelia and P. pentaurelia) on the basis of esterase zymograms. (ref. ID; 7704)
- Morphological features. Unfortunately, considering the great need of field ecologists and hydrobiologist for "quick-and-easy" cytological approaches to species identifications, the anatomical attributes of most of the species of the aurelia complex general do not provide data instantly usable in their certain taxonomic separation. In breif, nearly all of these 14 recently erected Paramecium species reveal features -after appliciation of some method of silver impregnation, for example- that neatly key them out to be simply members of the old Paramecium aurelia! This is of no help at all, of course. And as experienced field person can generally distinguish between species of the aurelia complex and other species of the genus using living material alone, without need of fixing, staining, etc. Nevertheless, careful analyses of light-microscopically visible morphological characters by multivariant biometrical methods have proven useful in some cases (e.g., see Gates and Berger 1976; Gates et al. 1975; Powelson et al. 1975). Morphogenetic features have yet to be explored in depth in the serice of taxonomy for the aurelia complex species, but their value in the higher-level systematics and phylogeny of ciliates is well established (Corliss 1968, 1979). Life cycle features, even cell size (in some cases), may be helpful to an extent; but a thorough knowledge of one's culture (this alone requires laboratory, not field, observations), employment of standard conditions, etc. would be required before the data could be used comparatively with a high degree of reliability. Ultrastructural information, so important in comparative ciliatology at higher taxonomic levels, is of practically no value at the subgeneric levels, for Paramecium or any other forms, because of its relative evolutionary conservativeness. (ref. ID; 7704)
- Ecological and biogeographical characteristics. There is inferential evidence (Sonneborn 1975) that certain of the 14 sibling species of the aurelia complex are limited to, or have preference for, restricted areas of the world (to give a negative example, strains of neither P. novaurelia nor P. quadecaurelia have been found to date in the United States). Yet few regions of the globe have been extensively sampled, so apparent distributional differences may not turn out to be true. On the other hand, the old belief that protist species, in general, are ubiquitous and thus have cosmopolitan ditributions is quite possibly not the case. However, our present knowledge is still too limited to draw a conclusion, one way or the other. "Morphological" species often appear to be found in very widely separated places; but, as in the case of Paramecium itself, such forms may sometimes be shown to be quite different genetically. Microhabitat. Differences have been little studied from a comparative point of view. Physiological adaptations in relation to species variability have also seldom been investigated in members of the aurelia complex. (ref. ID; 7704)
Other features of possible use. A variety of other physiological and "life cycle" or behavioral characters, raging from fission rate to temperature tolerance, might profitably be explored for use in separation of species of the aurelia complex. To date, such data are scarce and are generally considered unworthy of pursuit, since electrophoretic techniques are so reliable and relatively simple to carry out under standard laboratory conditions. Karyotypic and other (DNA, chromosome number etc.) nuclear characters, though little studied in extenso, seem to show as much or more variation within one of these species as between them. Mating reaction, mating type inheritance, etc., are, in the broadest sense, part of the "breeding relations" or genetics of these organisms and thus are valuable for purposes of species identification. However, their utilization comparatively, as menthioned above, requires living cultures of multiple strains testable under controlled conditions, a situation that rarely obtains for a general biologist. The use of intracellular symbionts (which we elect to call "xenosomes", using the name first applied by Soldo and Godoy, 1973, to similar bodies in certain marine scuticociliates), themselves now precisely indentifiable (Preer et al. 1974; Soldo 1974), remains an intriguing possibility for separation of members of the aurelia complex. But some stocks appear to carry no xenosomes; others need to be studied more intensively for their possible endosymbionts; and still others overlap in the kinds of microorganisms they carry. (ref. ID; 7704)
- Availability of strains (stocks): There are two major culture collections that maintain a variety of protozoan and algal species, the American Type Culture Collection (Rockville, Maryland) and the Culture Centre of Alage and Protozoa (Cambridge, England). The primary mission of both of these organization is to carefully maintain and make publicly available standardized or reference strains of these protists. Both collections maintain strains, also known as stocks, of species of the aurelia complex. The ATCC maintains all of its paramecium stocks as cryopreserved preparations, to insure long-term availability without change. A major portion of Sonneborn's collection was acquired by ATCC during the late 1970's, including "tester" stocks for all of the species. These are over 175 stocks at the ATCC now, an in the near future there will be more than 200 available. The number of paramecium strains maintained by the CCAP is not as extensive as at the ATCC, but there are some available there that are not obtainable from the ATCC. (ref. ID; 7704)
- Proposed preactical solutions: There are some practical considerations to be kept in mind in the designation of organisms belonging to the aurelia complex. It is of paramount importance that appropriate levels of identification be used if later comparison of data is to be possible. (ref. ID; 7704)
- For field biologists. The workers who are most likely to be faced with a practical problem of identification are field biologists. It is not possible (or at least with rare exception) to identify individual species of the aurelia complex solely on the basis of morphology. Although, as noted above, it is possible to make presumptive identifications using esterase zymograms, it is generally unlikely that most field biologists would be in a position to undertaken such an effort. However, it is now incorrect to use the specific disignation "Paramecium aurelia O.F.M, 1773" in their lists, tables, charts, etc. If the ecologist, limnologist, hydrobiolgist, plankton sampler (or whoever) is unable to do a detailed identification, then designation of the ciliate as "Paramecium sp." or as a "member of the P. aurelia complex" (perhaps using both with the second as a footnote to the first) is appropriate and ought to be employed. Such long-time authoritative field guides and textbooks as Kahl (1931) and Kudo (1966: and earlier editions), and now Bick (1972), Curds (1969), Dragesco (1970), Jahn et al. (1979) and others, are simply of no help in identification of these newly established (Sonneborn 1975) cryptic species of Paramecium. (ref. ID; 7704)
- For laboratory experimentalists. There is no excuse, in the case of laboratory workers, not to accurately identify the organism used in their researches. Without such presice identification of strains studied, the resulting data are almost usefuless. Reference stocks, the equivalent of reagent-grade chemicals (to which such careful attention is routinely paid), are readily available, as mentioned above. If a strain other than a reference stock is used, it is possible, in almost any modern laboratory, properly identify that organism to species-level, preferably by the enzyme analyses described earlier. (ref. ID 7704)
- For writers, teachers, and editors. The continued use of a legally discarded taxonomic name (in this case, Paramecium aurelia) by textbook writers, college professors and other teachers, and editors of journals is a disservice to science. Such carelessness perpetuates misinformation, and hence false biological information is continuously being presented as true to the entire scientific community. Such neglect of facts, wherther inavertent or otherwise, ignores important conclusions reached, often, only after years of research by competent investigators. Editors, the last to see a manuscript before it appears in print, bear a major responsibility with respect to the accuracy of a taxonomic name, the source of an experimental organism, and the precise strain or stock designations employed by an author. (ref. ID; 7704)
- Concluding remarks on the P. aurelia problem: There is no longer a species in the genus Paramecium named P. aurelia. Ciliates that appear to be the "old" species known by that name are probably assignable to the aurelia complex of sibling species. Appropriate breeding test and/or electrophoretic studies of their enzyme systems must be carried out to determine whether or not unknown strains or new isolates belong to one of the 14 cryptic or sibling (syngenic) species recognized by Sonneborn (1975) or represent a new species in this aurelia complex. Authenticated stocks are available as cryopreserved material, and it is advisable to use only such strains in experimental laboratory work if the resulting reseach data are to be meaningful. The population of "Paramecium aurelia" found in field work can be labeled only as "Paramecium sp." unless/until proper methods of precise identification have been carried out on them. An attractive possible alternative, using a "supraspecies" name (in effect, as a substitute for the words "a member of the aurelia complex"), is discussed in our Addendum. (ref. ID; 7704)
- Addendum: We are greatful to the Redacteur for permitting us to append a short section here concerning an excellent point brought to our attention by one of the reviewers of our manuscript, discussion of which however, would be difficult to incorporate directly to the pages of any preceeding sections. In effect, an alernative practical solution has been suggested for the problem of what name to use in identification of some sibling (jumelle) or syngenic species when sophisticated breeding or molecular techniques are not avaiable to the investgator (e.g., a field ecologist). The very recently proposed taxonomic-nomenclatural category of "supraspecies" -well described by its originators Genermont and Lamotte (1980), who acknowledge their indebtedness to nomenclaturist G. Bernardi- nicely links together taxonomically any group of species difficult to separate, one from the other, morphologically or ecologically. The term is not to be confused with the restrictive, little used, quite different category of "superspecies" (see Mayr 1969). Thus, instead of listing a population of organisms as "Paramecium sp." or "Tetrahymena sp.", indicating total ignorance of the precise (sibling) specific name to use and seemingly leaving only the genus as the first level of their identification, one could write: "Paramecium suprasp. aurelia" or "Tetrahymena suprasp. pyriformis". Usage of such terminology would clearly provide the reader with a "morphological type", as it were, but at an infrageneric (even infrasubgeneric) level. When/if the specific name is known, the new category could still be employed: for example, a biochemist might be working with Tetrahymena (suprasp. pyriformis) thermophila. In a way, the supraspecific category would replace the longer expressions "pyriformis complex" or "member of the pyriformis complex", the non-taxonomic notations recommended in a preceding section of this paper to field biologists, to be used as footnotes (in lists, tables, etc.) to the name Tetrahymena sp. One of the advantages of the idea is that it releases the term "complex" for informal usage at a still lower group level. Recall that Williams and Buhse (1983) have suggested that among the micronucleated members of the "T. pyriformis complex" one can recognize three well defined groups: the T. americanis complex, the T. borealis complex, and the T. thermophila complex. There appear to be very few disadvantages to this proposed practical solution, that is, using the supraspecies concept described above. Yet one, naturally, is that its very newness will, at first, make it unfamiliar and therefore possibly a little confusing to many readers. Also, the closeness of the spelling of the two words "supraspecies" and "superspecies" is unfortunate. From a taxonomic-nomenclatural point of view, however, we endorse with enthusiasm its future consideration by protologists. With respect to its usage with the two groups of ciliate sibling species under discussion in the present paper, only new serious problem arises. I is caused by the necessity (see Genermont and Lamotte 1980) of having a "nominate" species within each supraspecies; or, to put it another way, the Latin name of the supraspecies must be identical to the name of one (presumably the oldest) species included in the group. This causes no trouble in the case of Tetrahymena. T. (suprasp. pyriformis) pyriformis works very well. But for Paramecium, as discussed on earlier pages, there is no longer any taxonomic species P. aurelia and thus, legalistically, there can, at the moment, be no P. (suprasp. aurelia) aurelia! In their paper, our good French colleagues suggest rejection of Sonneborn's first binomen, P. primaurelia, replacing it with "P. aurelia", which would then allow a taxonomic-nomenclatural treatment parallel to that given for Tetrahymena above. But the matter cannot be handled so simply. Further discussion of this particular knotty and complex problem, however, is beyond the scope of the present paper. Its final proper resolution would involve, at the least, a carefully prepared lengthly pertition to the Intenational Commission on Zoological Nomenclature for future formal consideration by the commissioners. Unfortunately, at the present time, this restrict our giving complete support to the otherwsie attactive "alternative practical solution" discribed in this Addendum. (ref. ID; 7704)
- Paramecium aurelia Ehrenberg, 1838 (ref. ID; 1335, 1622, 1629, 2245) or 1848 (ref. ID; 3116) reported year? (ref. ID; 1618)
Syn; Paramecium aurelia Dujardin, 1841 - Paramecium aurelia Foissner et al., 1994 (ref. ID; 4488)
- Paramecium aurelia O.F. Muller, 1773 (ref. ID; 4611) or 1786 (ref. ID; 1622)
See; Paramecium caudatum - Paramecium aurelium Ehrenberg (ref. ID; 5624)
- Paramecium biaurelia (ref. ID; 3813, 3987, 4063, 4850)
See; Paramecium aurelia-complex - Paramecium bursaria (ref. ID; 191, 3292, 4903)
- Paramecium bursaria Ehrenberg (ref. ID; 1219, 1618) or (Ehrenberg, 1831) Focke, 1836 (ref. ID; 4611), (Ehrenberg) Focker, 1836 (ref. ID; 1335, 1622, 1629, 2245), (Ehrenberg) Focker, 1843 (ref. ID; 3116)
Syn; Loxodes bursaria Ehrenberg, 1831 (ref. ID; 4611) - Paramecium bursaria Fock, 1836 (ref. ID; 7399)
- Paramecium bursaria Focke (ref. ID; 3698)
- Paramecium calkinsi Woodruff, 1921 (ref. ID; 1335, 1622, 4611, 7399) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 4903)
- Paramecium caudatum Ehrenberg, 1833 (ref. ID; 4488, 4611) or 1838 (ref. ID; 1219, 1622, 1629, 2245, 3116) reported year? (ref. ID; 1618, 3342, 3698, 5462) reported author and year? (ref. ID; 191, 3292, 4064, 4903)
Syn; Paramecium aurelia O.F. Muller, 1786 (ref. ID; 1622) - Paramecium chilodonides (ref. ID; 1622, 4903)
- Paramecium chlorelligerum (ref. ID; 1620, 4903)
- Paramecium chrysalis O.F. Muller, 1786
See; Pleuronema crassum (ref. ID; 3116) - Paramecium colpoda Ehrenberg, 1831
See; Colpidium colpoda (ref. ID; 3116) - Paramecium compressum Ehrenberg, 1830 (ref. ID; 4730)
- Paramecium compressum Ehrenberg, 1838
See; Plagiotoma lumbrici (ref. ID; 4730) - Parameiucm cucllio Quenn., 1867 (ref. ID; 3116)
See; Plagiopyla nasuta (ref. ID; 3116) - Paramecium decaurelia (ref. ID; 4063)
See; Paramecium aurelia-complex - Paramecium dodecaurelia (ref. ID; 4063)
See; Paramecium aurelia-complex - Paramecium dragescoi (ref. ID; 4903)
- Paramecium duboscqui Chatton & Brachon, 1933 (ref. ID; 7399 redescribed paper) reported author and year? (ref. ID; 4830, 4903)
- Paramecium ficarium Kahl, 1928 (ref. ID; 1622, 4903)
- Paramecium glaucum Claparede & Lachmann, 1858 (ref. ID; 1622)
- Paramecium griseolum Perty, 1852(?)
See; Cryptochilum griseolum (ref. ID; 1622) - Paramecium jankowskii (ref. ID; 4903)
- Paramecium jenningsi Diller & Earl, 1958 (ref. ID; 1618, 4903) reported author and year? (ref. ID; 3813, 4064)
- Paramecium marinum Kent, 1881 (ref. ID; 1622)
Syn; Helicostoma oblongum Cohn, 1866 (ref. ID; 1622)
Paramecium multimicronucleatum-complex
- Paramecium multimicronucleatum syngen 1 (ref. ID; 3813, 4064)
- Paramecium multimicronucleatum syngen 2 (ref. ID; 3813, 4064)
- Paramecium multimicronucleatum syngen 3 (ref. ID; 3813, 4064)
- Paramecium multimicronucleatum syngen 4 (ref. ID; 3813, 4064)
- Paramecium multimicronucleatum syngen 5 (ref. ID; 3813, 4064)
- Paramecium multimicronucleatum Powers & Mitchell, 1910 (ref. ID; 1335, 1622) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 191, 3292, 3990, 4903)
- Paramecium nephridiatum V. Gelei, 1925 (ref. ID; 1622, 4903, 7486)
- Paramecium nigrum Burger, 1908
See; Frontonia atra Ehrenberg, 1833 (ref. ID; 1622) - Paramecium novaurelia (ref. ID; 4063, 4850)
See; Paramecium aurelia-complex - Paramecium octaurelia (ref. ID; 3813, 3899, 3987, 4063)
See; Paramecium aurelia-complex (ref. ID; 3987, 4063) - Paramecium pentaurelia (ref. ID; 4063)
See; Paramecium aurelia-complex - Paramecium polycaryum Woodruff, 1923 (ref. ID; 1335, 1622) reported year? (ref. ID; 1618) , Woodruff & Spencer, 1923 (ref. ID; 7399) reported author and year? (ref. ID; 191, 4903)
- Paramecium porculus (ref. ID; 4903)
- Paramecium primaurelia (ref. ID; 3813, 3829, 4063, 4760, 4850)
See; Paramecium aurelia subspecies 1 (ref. ID; 3829, 4063) - Parameicum pseudoputrinum Baumeister (ref. ID; 1219) reported author and year? (ref. ID; 4903)
- Paramecium putrinum Claparede & Lachmann, 1858 (ref. ID; 1622, 1629) or 1859 (ref. ID; 4611) reported year? (ref. ID; 1219, 1618) reported author and year? (ref. ID; 4903, 7599)
- Paramecium pyriforme Gourret & Roeser, 1886 (ref. ID; 1622)
- Paramecium quadecaurelia (ref. ID; 4063)
See; Paramecium aurelia-complex - Paramecium septaurelia (ref. ID; 3813, 4063)
See; Paramecium aurelia-complex - Paramecium sexaurelia (ref. ID; 3987, 4063)
See; Paramecium aurelia-complex (ref. ID; 3987) - Paramecium sonneborni Aufderheide, Daggett & Nerad, 1983 (ref. ID; 4061 original paper)
See; Paramecium aurelia-complex - Paramecium tetraurelia Sonneborn, 1975 (ref. ID; 4207) reported author and year? (ref. ID; 191, 3636, 3647, 3654, 3737, 3811, 3813, 3899, 4063, 4850, 6551, 7715, 7734, 7756)
See; Paramecium aurelia syngen 4 (ref. ID; 3737, 3811) - Paramecium traunsteineri (ref. ID; 1622)
- Paramecium tredecaurelia (ref. ID; 3987, 4063)
See; Paramecium aurelia-complex (ref. ID; 3987) - Paramecium triaurelia (ref. ID; 3813, 4063, 4850)
See; Paramecium aurelia-complex (ref. ID; 4063) - Paramecium trichium Stokes, 1885 (ref. ID; 1335, 1622, 2245, 7399) reported year? (ref. ID; 1219, 1618, 3342, 3698) reported author and year? (ref. ID; 191, 646, 7447)
- Paramecium undecaurelia (ref. ID; 4062)
See; Paramecium aurelia-complex - Paramecium woodruffi Wenrich, 1928 (ref. ID; 1335, 1618, 1622, 4611, 7399) reported author and year? (ref. ID; 4903)
Paramecium aurelia Ehrenberg, 1838 (ref. ID; 1335, 1622, 1629, 2245) or 1848 (ref. ID; 3116) reported year? (ref. ID; 1618)
Synonym
Paramecium aurelia Dujardin, 1841Descriptions
Two small vesicular micronuclei, a massive macronucleus; two contractile vacuoles on aboral surface; posterior end more rounded that P. caudatum; in fresh water. (ref. ID; 1618)Measurements
120-180 um long. (ref. ID; 1618)Paramecium bursaria Ehrenberg (ref. ID; 1219, 1618) or (Ehrenberg, 1831) Focke, 1836 (ref. ID; 4611), (Ehrenberg) Focker, 1836 (ref. ID; 1335, 1622, 1629, 2245), (Ehrenberg) Focker, 1843 (ref. ID; 3116) reported author and year? (ref. ID; 3292)
Synonym
Loxodes bursaria Ehrenberg, 1831 (ref. ID; 4611)Descriptions
Foot-shaped, more or less flattened; uniform ciliation except for a group of long caudal cilia; green with symbiotic zoochlorellae; a long broad vestibulum leads to the buccal cavity, the buccal ciliary apparatus is characterized by 2 "peniculi"; 1 compact micronucleus; 2 contractile vacuoles; numerous prominent trichocysts. (ref. ID; 1219)Foot-shaped, somewhat compressed; green with zoochlorellae as symbionts; a compact micronucleus; a macronucleus; two contractile vacuoles; in fresh water. (ref. ID; 1618)
Measurements
Length 180-300 um. (ref. ID; 1219)About 100-150 by 50-60 um. (ref. ID; 1618)
Paramecium calkinsi Woodruff, 1921 (ref. ID; 1335, 1622, 4611, 7399) reported year? (ref. ID; 1618)
Descriptions
Foot-shaped; posterior end broadly rounded; two vesicular micronuclei; two contractile vacuoles; in fresh, brackish and salt water. (ref. ID; 1618)Measurements
100-150 by 50 um. (ref. ID; 1618)Paramecium caudatum Ehrenberg, 1833 (ref. ID; 4488, 4611) or 1838 (ref. ID; 1219, 1622, 1629, 2245, 3116) reported year? (ref. ID; 1618, 3342, 3698, 5462) reported author and year? (ref. ID; 3292, 4064)
Synonym
Paramecium aurelia O.F. Muller, 1786 (ref. ID; 1622)Descriptions
Cigar-shaped, posterior end bluntly pointed and with a group of long cilia, ciliation otherwise uniform; vestibulum long and slightly oblique; buccal cavity with one endoral membrane and 2 peniculi; 1 ellipsoid macronucleus and 1 compact micronucleus; 2 contractile vacuoles, each with radial canals, near the aboral surface, numerous trichocysts, which may discharge explosively, all over the body. (ref. ID; 1219)With a compact micronucleus, a massive macronucleus; two contractile vacuoles on aboral surface; posterior end bluntly pointed; in fresh water. The most widely distributed species. (ref. ID; 1618)
Measurements
Length 90-150 um. (ref. ID; 1219)180-300 um long. (ref. ID; 1618)
180-260x40-50 um. (ref. ID; 3342)
Paramecium duboscqui Chatton & Brachon, 1933 (ref. ID; 7399 redescribed paper) reported author and year? (ref. ID; 4830, 4903)
Descriptions
[ref. ID; 7399]The body shape with its rounded posterior end and broad anterior part is of the "bursaria type" (Wichterman 1986). The oral groove is broad and oblique, and the anterior-right part of the body extends and twists to the ventral side, so that the whole body is kidney-shaped in lateral view when the animal is moderately starved. The body surface is uniformly ciliated except for the three or four long caudal cilia at the posterior end. The body length of 30 randomly chosen, fixed specimens ranged from 80 to 150 um. (ref. ID; 7399)
- Argentophilic structures: In silver-impregnated specimens, the pellicular pattern forms hexagons over most of the body surface. Rhomboids are restricted to the left side of the preoral suture, and quadrilaterals are restricted to both sides of the postoral suture and the right side of the preoral suture. The preoral suture, an argyrophilic line, forms a bow-shaped curve to the organisms left. The cytoproct occupies the posterior half of the postoral suture. Two contractile vacuole pores, one for each contractile vacuole, are visible on the dorsal surface. (ref. ID; 7399)
- Buccal apparatus: The vestibulum is shallow and ends in the middle or a little above the middle of the body at the buccal overture. The endoral membrane lies along the entire right edge of the buccal overture, but from its origin to its middle, the endoral dikinetids are zig-zag or single file. From the middle to the end, the kinetosomes of the dikinetids are side by side. The anarchic field is clearly visible at the right of the posterior end of the endoral membrane. Two peniculi lie on the left wall of the buccal cavity; each contains four rows of kinetosomes. The posterior end of the dorsal peniculus does not turn to the ventral and right side although that of the quadrules does. (ref. ID; 7399)
- Stomatogenesis: During cell fission, as in other species of Paramecium, the buccal pouch for the future opisthe originates from the anarchic field and the inner portion of the right wall of the vestibulum. Three nascent membranelles extend out of the vestibulum and lie on the somatic pellicle at the right of the postoral suture. After the new buccal cavity separates from the old, the two buccal apparatuses are juxtaposed. This stomatogenesis strongly resembles that of P. bursaria (Shi 1980). (ref. ID; 7399)
- Nuclear morphology: One ellipsoid macronucleus is present just above the center of the cell. In protargol-stained interphase cells, the nucleoli in the macronucleus often aggregates into a coarse network. This network is identified as nucleolar because it stains as nucleoli do in other ciliate species not as microtubules or other supporting structures do. The number of micronuclei range from zero to six, but two micronuclei are usual. Of 1,000 cells from 10 stocks examined, 685 (68%) had two micronuclei. Micronuclei in interphase are vesicular and shaped like long spindles (ca. 5.1 um x 1.2 um in hematoxylin preparations and ca. 10.2 um x 3.2 um in protargol preparations. (ref. ID; 7399)
- Mating types: By mixing evey two combinations among 46 stocks collected from the same place, only two mating types have so far been detected. They are designated as syngen 1 in this species. This is the first report of conjugation in this species. Whether there were other mating types and syngens in collections from other locations remain to be studied. (ref. ID; 7399)
- Trichocysts: The subpellicular trichocysts are perpendicular to the surface and appear more conspicuous than in other species. In protargol preparations, the main body of the trichocyst is rather thick, and its width is nearly equal to one half of its length. The length of the tip is about two-fifths that of the main body. (ref. ID; 7399)
- Contractile vacuoles: Both contractile vacuoles are vesicle-fed. Immediately after systole the vesicles expand. At diastole, fibrils supporting the walls of the contractile vacuole are visible in the living ciliate. Probably these correspond to the tracts of microtubules that spiral around the wall of the contractile vacuole in other species of Paramecium. (ref. ID; 7399)
- Swimming behavior: In contrast to all other species of Paramecium except P. calkinsi, this ciliate characteristically spirals to the right on its long axis (clockwise) when swimming. There are two spiraling modes in swimming, and extremely narrow spiral at greatest speeds and a long spiral at average speeds. These figures indicate that the forward progress of P. duboscqui when it is swimming fastest may be slower than its forward progress when it is swimming at slower speeds. (ref. ID; 7399)
Remarks
Chatton and Brachon (1933) reported that they collected their strain from a sewer in Banyuls-sur-Mer in January and April in 1933. The collecting seasons and the habitats of the two strains are so similar that the phenomenon cannot be considered as accidental. The temperature of sewer water in winter and early spring in France should be at its lowest for the year. It thus seems clear that the French strain of P. duboscqui must also be adapted to lower temperatures. At least two species of Paramecium of the bursalia group, Paramecium polycaryum and Paramecium woodruffi, are rarely found, although P. polycaryum is worldwide in distribution. Paramecium duboscqui may not be a rare species at all. Its absence from collections may reflect the fact that fewer collections of protozoa are made in the wintertime. Paramecium duboscqui clearly belongs to the bursaria group based on its nearly body-shape, i.e. the truncated anterior end, the rounded posterior end, the dorso-ventral flattening, the terminal cytoproct, and the pattern of stomatogenesis (Shi 1980). Its characteristics led the authors to the conclusion that Woodruff (1921) is fully correct in his classification of the species of Paramecium into "aurelia" and "bursaria" groups. They are really two natural groups formed in the long proccess of phylogeny in Paramecium. The twist of the anterior body of P. duboscqui and its reniform outline in lateral view are unique among the species of the bursaria group. Swimming in a right spiral is a characteristic shared only by P. calkinsi. In its contractile vacuole structure, P. duboscqui resembles P. trichium, which also has vesicle-fed contractile vacuoles. These are the only species of Paramecium that lack collecting canals. Paramecium trichium also has an unusual nucleolar structure, not like that of P. duboscqui, but nucleoli "pocketed" in the surface of the macronucleus. This species is the smallest in the genus, but some strains are large enough to overlap with P. duboscqui size range. It has a subcylindrical body, a single micronucleus, and a complex mating system. The morphological and breeding differences between these two species are significant enough to separated them. The right spiral swimming of P. calkinsi is a characteristic shared by P. duboscqui, but P. calkinsi has a buccal overture below the center of the body, compared to P. duboscqui buccal overture at or above the midline. It is euryhaline and can be found in freshwater ponds as well as marine tide pools. The breeding system of P. duboscqui, one syngen and two mating types, is like those of P. polycaryum and P. woodruffi, but more collections and breeding studies of these three species may well turn up other syngens as it did in P. calkinsi. It seems unlikely, however, that the breeding systems in these species will ever approach the complexity of the breeding systems of P. bursaria and P. trichium with their many syngens and multiple mating types. (ref. ID; 7399)Temperature restrictions
Many collections from Majia Ditch were made year round for 14 years. Winter in Harbin is from November to March, and the water temperature in the shallow water at the edge of the ditch drops below 10 degrees C. Paramecium duboscqui was collected only at these temperatures. It was collected frequently during winter, but when water temperature rose to 15 degrees C, it disappeared. Where it summers is unknown, and even in winter, it did not appear in collections from other sites. (ref. ID; 7399)Type locality
Collected from running water in Majia Ditch, Harbin, China, the overall length of which is more than 30 km. Industrially polluted water and domestic sewage drain into its upper reaches, and it empties into the Songhua Riber in Harbin city proper. (ref. ID; 7399)Type materials
The slides of the type-specimens and the living strains are preserved in the Laboratory of Protozoology in Harbin Normal University. (ref. ID; 7399)Paramecium jenningsi Diller & Earl, 1958 (ref. ID; 1618) reported author and year? (ref. ID; 3813, 4064)
Descriptions
Resembles P. aurelia in general morphology; two micronuclei larger than those of P. aurelia; macronuclear anlagen with long persisting chromatinic centers. (ref. ID; 1618)Measurements
115-218 um long. (ref. ID; 1618)Paramecium multimicronucleatum< Powers & Mitchell, 1910 (ref. ID; 1335, 1622) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 3292, 3990)
Descriptions
3-7 contractile vacuoles; four or more vesicular micronuclei; a single macronucleus; in fresh water. (ref. ID; 1618)The somatic kinetosomes, single or double, appear as small dark granules; issuing from them is a kinetodesmal (Kd) fiber that runs anteriorly and to the right of the kinety, overlapping with more anteriorly located fibers to form a single kinetosomes are more closely spaced and the kinetodesmata are thicker by virtue of being formed by a larger number of overlapping Kd fibers. (ref. ID; 3990)
Measurements
The largest species, 200-330 um long. (ref. ID; 1618)Paramecium nephridiatum V. Gelei, 1925 (ref. ID; 1622, 4903, 7486)
Descriptions
The cell shape with its rounded posterior end, and broad anterior part is of the "bursaria" type (Wichterman 1986) or "woodruffi" type according to Jankowski (1969). The oral opening is situated a little anterior to the middle of the cell. The cell surface is uniformly ciliated except for several long caudal cilia located close to the posterior end of the dorsal side, but not on the top. The abundant subpellicular trichocysts are distributed uniformly. Specimens were about 145x47 um long in vivo but shrank 10% after the silver nitrate impregnation. In silver-impregnated specimens, there were ~38 rows of ventral kineties and ~35 dorsal kineties. The preoral suture is distinct, but the postoral suture is very obscure. The cytoproct is situated in the posterior third. The vestibular zone is conspicuous and is terminated by the distinctive shape of the buccal opening. Two peniculi and open quadrulus are located on the dorsal and left wall of the buccal cavity. The endoral membrane is situated along the entire right edge of the buccal opening but its dikinetids are not recognizable in all specimens. It has 15 dikinetids on the average. The buccal cavity size various around 30 um. On the dorsal side of the body the contractile vacuole pores are very distinctive both on impregnated specimens as well as in living cells. Usually, both contractile vacuoles have more than one pore each, typically two or three. However, we have found two stocks (WCh-1 and WS-12) where one of the contractile vacuoles quite often (up to 50%) had only one pore. Both contractile vacuoles usually have 8-14 collecting canals, ten on average. Numerous crystals were very often found in the cytoplasm, but their quantity and location varied, probably depending on the culture conditions. During cell division, the anterior daughter cell (proter) more closely resembled the maternal cell than the opisthe which, at first, looks highly similar to some other peniculines. The nuclear apparatus is located in the anterior part of the cell. One slightly ellipsoid or ovoid macronucleus, ~30x36 um in living cell and ~17x23 um in stained cells, on average, resides just anterior to the equator of the cell. In Feulgen-stained cells the macronucleus has a very intense colour. The three to four spherical micronuclei of the "endosomal" type (Fokin 1997), ~3 um in diam. (on average) are distributed irregularly along the anterior part of cell. Endocytobiotic bacteria are often found in the cytoplasm (Fokin 1989) and can also be found in the perinuclear space and in the macronucleus (Fokin 1989; Fokin and Sabaneyeva 1997). The species is characterized by a binary mating type system. Using more than 20 stocks from different localities, two unambiguous mating types have been detected. The old macronucleus fragments before the separation of the conjugants. The new macronuclear anlagen are four in number. Selfing in stock cultures takes place on rare occasions. (ref. ID; 7486)Swimming behavior
During swimming this species spirals on its long axis in both directions (Fokin 1987). We could no find any simple triggers (food, time) for changing this swimming direction. Typically, "left spiral swimmers" and "right spiral swimmers" were present at the same time in the culture. During the several years of investigation of this trait there was some preference of the cells from the same stocks to spiral in the left direction. (ref. ID; 7486)Remarks
Gelei (1925) described a new species of Paramecium, Paramecium nephridiatum, based on the material which he had found in his laboratory aquarium. This population was in fact a mixture of the new species and Paramecium caudatum (Gelei 1938). This was the reason why some features of the new ciliate were similar to P. caudatum so that no one recognized this new species in nature, though reference to P. nephridiatum was made by Kahl (1931) and Kalmus (1931). Gelei (1938) redescribed the species from a native population (Tisza River, Szeged, Hungary) using a "clean culture". For unknown reasons, this new description did not attract the attention of protozoologists and in all subsequent reviews, P. nephridiatum was considered a nonvalid species (Vivier 1974; Wichterman 1953, 1986) even when the article of Gelei (1938) was listed in the references. Only once was P. nephridiatum mentioned in a short abstract as a species living in Florida, USA (Bovee 1983), although characters of the species were not listed in this publication. Since 1983, one of us (S. F) has repeatedly collected a species of Paramecium with multiple contactile vacuole pores, which is a distinctive trait of P. nephridiatum, although it was considered for a time as a feature of Paramecium woodruffi (Agamaliev 1983; Fokin 1986), Jankowski (pers. commun.). (ref. ID; 7486)Occurrence and ecology
A number of stocks of P. nephridiatum were isolated from the sea shores of northern Europe: the North, Baltic, White, and Barents Sea coasts. It was detected during sampling in Woods Hole, MA, USA, Atlantic Ocean and on Sakhalin Island, Sea of Okhotsk. The salinity of these samples varied from 1.5-32 0/00. The species was also found in a fresh-water body in Jerusalem Zoo, Israel. Samples were taken mainly during the summer, from mid-April (Wood Hole) to November (North Sea coast). Sampling of the same wild population of P. nephridiatum (Sredny Island, White Sea, Russia) has been repeated every year since 1990 to observe long-time changes in the population, as well as the euryhaline ability of the species. This population, as well as the euryhaline abiliy of the species. This population was present at salinities from 4-35 0/00 and in the temperature range from 10-25 degrees C. Very often the populations of P. nephridiatum occurred at the lower limit of oxgen concentrations. They were mainly feeding on bacteria. In the same samples these other ciliates were usually found: Prorodon sp., Frontonia marina, Metopus sp., P. calkinsi, P. woodruffi, and sometimes P. duboscqui. (ref. ID; 7486)Paramecium polycaryum Woodruff, 1923 (ref. ID; 1335, 1622) reported year? (ref. ID; 1618), Woodruff & Spencer, 1923 (ref. ID; 7399) reported author and year? (ref. ID; 191, 4903)
Descriptions
Form similar to P. bursaria; two contractile vacuoles; 3-8 vesicular micronuclei; in fresh water. (ref. ID; 1618)Measurements
70-110 um long. (ref. ID; 1618)Paramecium putrinum Claparede & Lachmann, 1858 (ref. ID; 1622, 1629) or 1859 (ref. ID; 4611) reported year? (ref. ID; 1219, 1618) reported author and year? (ref. ID; 4903, 7599)
Descriptions
Similar to P. bursaria but without zoochlorellae; macronucleus elongated, kidney-shaped; 1 micronucleus; only 1 contractile vacuole; no trichocysts (Lepsi, 1926; Kahl, 1930) Records of P. putrinum are scanty, and it is in fact doubtful whether the species is valid. This problem is of some saprobiological interest since Liebmann (1962) classified P. putrinum as a polysaprobic indicator organism. On the other hand, Sramek-Husek (1954) failed to find any specimens of P. putrinum during his saprobiological studies in Czehoslovakia; all representatives of the genus Paramecium that he found in the more heavily polluted zones were referable to P. trichium. Therefore, Sramek-Husek submitted that the polysaprobic "P. putrinum" of earlier authors is really P. trichium Stokes. P. putrinum sensu Liebmann has 2 contractile vacuoles, both shown with radial canals; P. putrinum sensu Liebmann was recorded from the sludge of polluted reservoirs, dammed rivers, lakes etc., in the upper zone of trickling filters, sewage drains, and in floating tufts of Sphaerotilus. Taxonomic studies of "P. putrinum" and P. trichium are thus urgently required. (ref. ID; 1219)Similar to P. bursaria, but a single contractile vacuole and an elongated macronucleus; no zoochlorellae; in fresh water. (ref. ID; 1618)
The ultrastructural study of postoral microtubules. (ref. ID; 7599)
Measurements
About 130 um long (ref. ID; 1219)80-150 um long. (ref. ID; 1618)
Paramecium sonneborni Aufderheide, Daggett & Nerad, 1983 (ref. ID; 4061 original paper)
See
Paramecium aurelia-complexDescriptions
[ref. ID; 4061]Etymology
The new species has been named Paramecium sonneborni n. sp. in honor of the late Dr. Tracy M. Sonneborn of Indiana University. (ref. ID; 4061)Type locality
College Station, Texas (Lat. 30 degrees 37'55''N, Long. 96 degrees 18'15''W). (ref. ID; 4061)Type-specimens
Holotype and paratype slides (Nos. USNM31926 and 31927) have been deposited in the Ciliate Type Slide Collection, Smithsonian Institution, Washington, D.C. Paratype specimens remain in the collection of the senior author. (ref. ID; 4061)Type culture
Paramecium sonneborni stock TAMU:0580:1 ATCC 30995) (ref. ID; 4061)Measurements
Maximum body length of 26 fixed, silver-impregnated specimens ranged from 130 to 186 um, with a mean length of 154.4 um (+/-2.9 SE). Maximum body width of the 26 cells ranged from 39 to 64 um, with a mean width of 50.3 um (+/-1.3 SE). (ref. ID; 4061)Paramecium tetraurelia Sonneborn, 1975 (ref. ID; 4207) reported author and year? (ref. ID; 191, 3636, 3647, 3654, 3737, 3811, 3813, 3899, 4063, 4850, 6551, 7715, 7734, 7756)
See
Paramecium aurelia syngen 4 (ref. ID; 3737, 3811)Descriptions
Parallel study of stomatogenic and micronuclear activities in sexual reproduction. (ref. ID; 7715)Paramecium trichium Stokes, 1885 (ref. ID; 1335, 1622, 2245, 7399) reported year? (ref. ID; 1219, 1618, 3342, 3698) reported author and year? (ref. ID; 191, 646, 7447)
Descriptions
Body shape highly variable, more or less ellipsoid, somewhat flattened; ciliation uniform except for a group of long caudal cilia; vestibulum and buccal equipment similar to those of P. bursaria; 1 macronucleus; 1 compact micronucleus; 2 contractile vacuole, each with a convoluted outlet; unlike other members of its genus, P. trichium has a contractile vacuole without radial canals but with tributary vacuoles; numerous trichocysts all over the body; posterior part of the body more or less filled with dark granules. According to Sramek-Husek (1954), P. pseudoputrinum Baumeister is referable to P. trichium. (ref. ID; 1219)Oblong, somewhat compressed; a compact micronucleus; two contractile vacuoles deeply situated, each with a convoluted outlet; in fresh water. (ref. ID; 1618) [ref. ID; 7447]
- Morphology of the non-dividing (morphostatic) cells
Throughout this paper, the directions right and left will be described according to the definition of Ng and Frankel (1977) as ".., right and left refer to the observer's right and left, assuming that he stands inside the animal so that his anterior-posterior axis coincides with that of the animal, and keeps turning around his longitudinal axis to face the surface of the animal," unless otherwise indicated. (ref. ID; 7447)- Basal bodies. In the anterior 3/4 to 4/5 of the cell surface, almost all cortical units possessed two closely spaced basal bodies (2BB), while each unit of the remaining region included one basal body (1BB). These units, hereafter, will be called 2BB units and 1BB units, respectively. In the 1BB unit region, paired basal bodies were not usually observed, while 1BB unit were often sporadically distributed over the 2BB unit region. Each unit usually included one parasomal sac (Ps), although in several units Ps could not be detected by the Chatton-Lwoff method. (ref. ID; 7447)
- Kinetodesmal fibers. Kinetodesmal (Kd) fibers arose close to the single basal body in 1BB units and to the posterior one of the paired basal bodies in 2BB units, and always extended forward and along the right side of the kinety. The Kd fiber of the 2BB unit was approximately 3-5 um in length, while their length in 1BB units tended to be slightly shorter than in 2BB units. (ref. ID; 7447)
- Kineties. When a paramecium is observed from its anterior pole, kineties are numbered clockwise with reference to the postoral suture (PO). Kinety No. 1 (K1) has a few cortical units and extends rearward along the right side of PO from the posterior margin of the buccal cavity (BC); and enumeration of kineties proceeds to the cell's right (viewer's left). Thus, the kinety on the left side of PO is No. n (Kn), with n taking the value of the total number of kineties. There are two or three rows of single basal bodies in the slender narrow (SN) region lying between kinety No. 3 (K3) and the paroral membrane (PM), which is a file of 16 to 25 closely packed paired basal bodies. However, we could not determine whether they consisted of the same components as the other cortical units, and therefore were anterior portions of kinety No. 1 and 2, or not. Furthermore, there were two short kineties just anterior to the SN region. The left one of these included two or three 2BB units and the other included three or six 2BB units. We also could not determine whether these were K1 and K2. In some species of paramecia, a permanent anarchic field has been described in the comparable area (Frankel 1989; Hufnagel 1969; Patterson 1981). However, we could not find such a field in P. trichium. Twenty specimens is which almost all basal bodies could be traced in eight representative kineties from Kn-9 to Kn-16 were obtained by the Chatton-Lwoff method. The number of 2BB and 1BB units was examined in these representative eight kineties. In these kineties, the mean number (+/-SD) of 2BB, 1BB, and total units per kinety was 27.60+/-3.45 (range 18-38), 6.67+/-1.83 (3-12), and 34.27+/-3.85 (25-43) (n=160 kineties) respectively, although each kinety tended to contain slightly more units that its right-hand neighbor. (ref. ID; 7447)
- Polygonal cortical ridges. When deciliated paramecia were fixed with Parducz's fixative and were observed by scanning electron microscopy, their cell surfaces were seen to be covered with a regularly arranged meshwork of polygonal cortical ridges. Since a territory bounded by the polygonal cortical ridges was regarded as a cortical unit, longitudinal rows of these territories were considered to correspond to kineties. In these deciliated paramecia, one or two remnant stub(s) of cilium(a) existed in each depression area of the cortical units. In the ventral surface of a deciliated cell, the oral groove region was covered with cortical units containing two remnant stubs of cilia (2C), although the most anterior cortical unit of each kinety on the right side of preoral suture (PR) possessed only one remnant stub of cilium (1C). Since this region consisted of 2BB units, the units of the region will be called two cilia-bearing-basal-body units (2C-2BB unit). In contrast, the remainder of the cell surface was covered with cortical units containing only one remnant stub of cilium (1C). The posterior 1/4 to 1/5 of the cell surface was covered with 1BB units and the remainder consisted of 2BB units, so that these units could be named 1-cilium-1-basal-body (1C-1BB) just and 1-cilium-2-basal body (1C-2BB) units, respectively. Also, several non-ciliated cortical units were sporadically distributed in the one cilium unit region. In the 1C-2BB unit region, two-cilia units were sometimes observed in non-deciliated cells. For comparison with the results from the analyses of silver impregnated specimens, the number of two-cilia units, one-cilium units, no-cilium units, and total units per kinety were examined in eight representative kineties from Kn-9 to Kn-16. In these kineties, the mean number (+/-SD) of two-cilia, one-cilium, no-cilium, and total cortical units per kinety was 8.71+/-1.52 (range 5-13), 27.31+/-2.62 (20-34), 0.14+/-0.62 (0-4) and 36.16+/-2.32 (29-42) (n=160 kineties), respectively. (ref. ID; 7447)
- Reorganization of cortical structures during cell division
The reorganization of cortical structures during cell division could be classified for convenience into five stages according to morphological characters.- Stage 1. When the cell surface was observed by scanning electron microscopy, appearance of a bare area was the first recognizable sign of morphogenetic events prior to cytokinesis. This unciliated, relatively flattened, area is located just posterior to the oral apparatus. This area formed as a result of the disappearance of cilia and polygonal cortical ridges in the posterior parts of kineties from K1 to K6, although we could not determine whether the cilia are resorbed into the cytoplasm or detached from the cell surface. On the other hand, in cells stained by the Chatton-Lwoff method, proliferation of basal bodies in the posterior part of SN region was the first visible morphogenetic event. Subsequently, in the posterior portion of K3 to K6, boundaries of cortical units gradually became obscure, and basal bodies rapidly proliferated. These basal bodies formed a long slender anarchic field parallel to the longitudinal axis of the cell. However, we could not determine whether these basal bodies of the anarchic field bore cilia or not, and also could not clarify the timing and spatial relationships between the sequential morphogenetic events visualized by the silver impregnation technique and the appearance of the bare area observed by scanning electron microscope. (ref. ID; 7447)
- Stage 2. In the fully extended anarchic field, basal bodies were progressively arranged in the three files from the posterior end to the anterior end of the field as well as from the right side to the left side of the field. Each of the files was made up to four longitudinal rows of basal bodies. Of these, the right-most file was a primordium of the quadrulus and the remaining two were those of peniculi of the prospective posterior daughter cell. Since the deciliation technique used in this study could not detach the cilia of the anarchic field, it was found by scanning electron microscopic observation of deciliated cells that most, if not all, basal bodies of the oral primordium bore cilia at this stage. The beginning of fission line formation is another remarkable morphogenetic event of this stage. In each of several 1C-2BB units located along the equatorial right ventral region near the oral primordium, pre-existing Kd fibers were shortened and a new basal body was added just anterior to the paired basal bodies. In some species of paramecia, it has been reported that all old Kd fibers are resorbed and entirely new ones are formed prior to cytokineties (Fernandez-Galiano 1978, Iftode et al. 1989). However, in P. trichium we could not observe such a resorption of old Kd fibers. The fission line began to be formed as a discontinuity between such morphologically changed cortical units and unchanged units anterior to these altered units in each of kineties. The zone of discontinuity appeared at the right anterior region of the oral primordium. Then fission line formation rapidly progressed clockwise when the paramecium was observed from its apical pole. With the progression of fission line formation, a shortening of the preexisting Kd fiber and an addition of a basal body immediately anterior to the paired basal bodies occurred in cortical units that were located in the 1C-2BB region posterior to the fission line. Late in this stage, the triplet basal bodies of each cortical unit began to separate from each other and a new short Kd fiber appeared in each of the anterior two basal bodies of the unit. These sequential morphogenetic events progressed toward the posterior from the fission line as well as to the right side from the PO. By the end of this stage, the posterior margin of this morphogenetically changing area reached approximately to the middle of the 1C-2BB region, located to the right of the PO. In the remainder of the cell surface, visible morphogenetic changes had not yet occurred at the end of this stage. (ref. ID; 7447)
- Stage 3. The formation of a second fission line was one of the most remarkable aspects in this stage. The second fission line appeared as a discontinuity in each kinety, formed in the same manner as the first fission line, along the equatorial zone of the cell surface region to the left of the oral primordium. The disruption of kineties sequentially propagated to the left from the Kn: the formation of this fission line progressed counterclockwise when the paramecium was observed from the apical pole of the cell. This fission line encountered the first one on the dorsal surface, and they then joined to become a single fission line. As a result, the region anterior to the fission line became the cell surface of the prospective anterior daughter cell, while the region posterior to the fission line became the prospective posterior daughter cell. Gillies and Hanson (1968) noted that "the fission line appears at the (cell's) left posterior end of the (oral) anlage and during stage III develops around the cell, passing dorsally and then returning ventrally to the (cell's) right anterior side of the anlage." This mode of fission line formation was never observed in our paramecia. To determine whether preexisting cortical units were equally distributed to both presumptive daughter cell surfaces by the formation of the fission line, in Chatton-Lwoff silver-impregnated stage-3 cells the number of cortical units per kinety was examined in eight representative kineties from Kn-9 to Kn-16, comparable with those of vegetative morphostatic cells. The posterior daughter cell surface received, significantly, one or two more units than the anterior surface (t-test, p<0.02). (ref. ID; 7447)
- Stage 4. Formation of the new buccal cavity for the prospective posterior daughter cell and constriction of the fission furrow along the fission line began to occur at this stage. In the prospective posterior daughter cell, differentiation of three oral membranelles, the two peniculi and a quadrulus, for the new oral apparatus had been completed by this stage and these ciliated structures were arranged on the cell surface to the posterior right of the old mouth parts. Then, a slit-like bare area appeared between the right most membranelle and an anarchic strip that consisted of many basal bodies randomly and closely distributed, located the right side of the membranelles. Subsequently, this bare are began to invaginate from the cell surface while extending in area. With the development of the buccal cavity (BC), the three membranelles sank and migrated to their final position along the left wall of the cavity. A new paroral membrane (PM) was differentiated at the left edge of the anarchic strip during the final stage of the BC formation. Another remarkable morphogenetic property of this stage was the appearance of cytospindle-like structures (Cohen and Beisson 1988; Cohen et al. 1982; Delgado et al. 1990; Fleury and Laurent 1995; Frankel 1989; Iftode et al. 1989; Jerka-Dziadosz et al. 1992), which could only be visualized by the Fernandez-Galiano method. They appeared as discontinuous wavy lines in the spaces between kineties early during this stage. Then, the developed to continuous wavy lines by the end of this stage. (ref. ID; 7447)
- Stage 5. The morphogenetic events of the prospective posterior daughter cell surface were almost completed except for a new PR formation by the start of this stage. In the prospective anterior daughter cell surface the remaining morphogenetic events progressed rapidly from the fission furrow to the anterior pole of the cell and were completed by the end of this stage. Furthermore, in the 2C-2BB unit region the Kd fiber of each unit extended again to its final length without either proliferation of units or new cortical-component formation. With the progression of these morphogenetic events, the contractile ring rapidly contracted and finally divided the mother cell to two newborn daughter cells. (ref. ID; 7447)
Examined material
Clone 4B-1 (mating type IV) of the ciliate Paramecium trichium. This clone, which was esatblihsed in January 1991, is one of the F1 progeny of a crossbetween stocks OM4(IV) and MMB(III), which were kinely provided by Dr. Toshikazu Kosaka, Hirosima University. The clone 4B-1 was sexually mature and did not show any sign of senility during the course of this work. (ref. ID; 7447)Measurements
Length 50-120 um. (ref. ID; 1219)50-105 (80-90) um long. (ref. ID; 1618)
80-110x45-60 um. (ref. ID; 3342)