Euplotes
Euplotes Ehrenberg, 1830 (ref. ID; 662, 4905, 7354) or 1831 (ref. ID; 2014, 7513)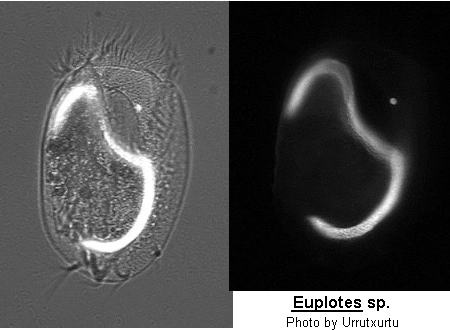
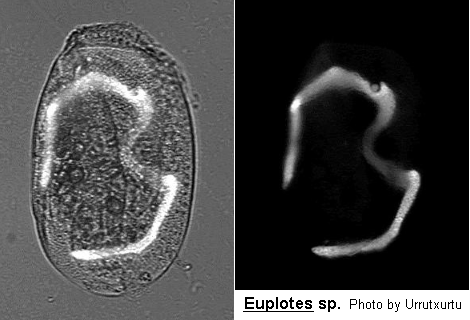
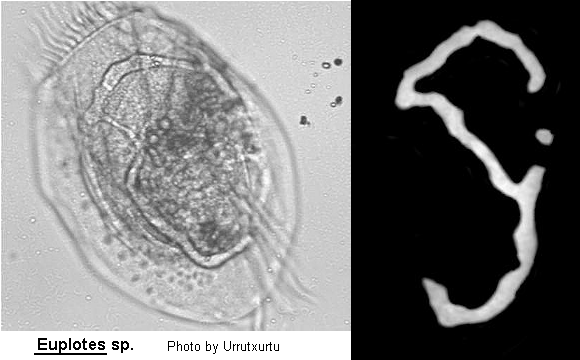
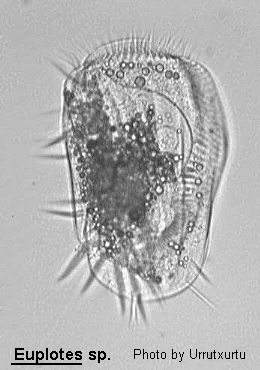
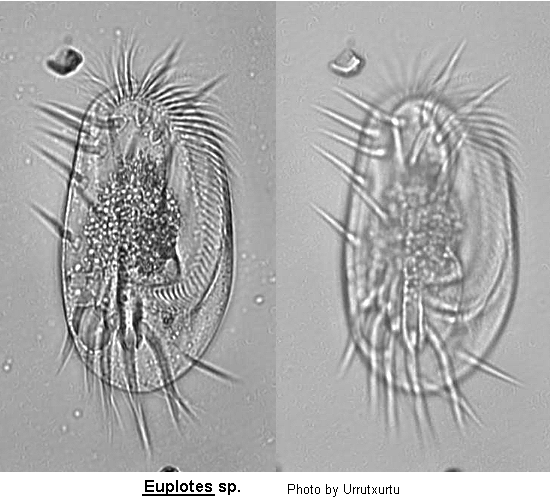
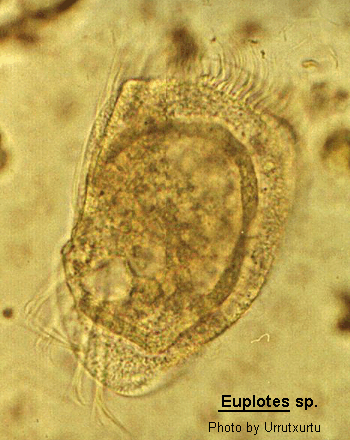
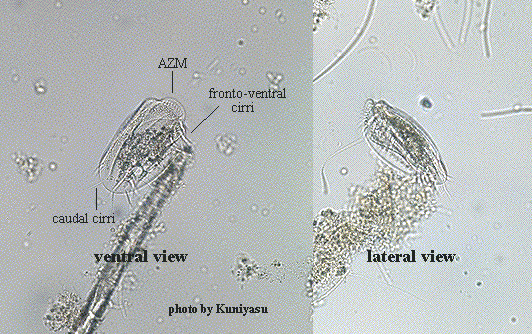
Class Polyhymenophora Jankowski, 1967: Order Hypotrichida Stein, 1859 (ref. ID; 4893)
Order Euplotida: Family Euplotidae (ref. ID; 7354) Synonym Crateromorpha Perejaslawzewa, 1886 (ref. ID; 2014); Euploea Ehrenberg, 1830 (ref. ID; 2014); Ploesconia Bory, 1825 (ref. ID; 2014) [ref. ID; 2014]
Inflexible, dorso-ventrally flattened, dorsally ridged, oval body with very large AZM usually extending at least two-thirds the body length, broadly triangular and often supported anteriorly by a cytoplasmic collar. Marginal rows of cirri absent but usually 1 or 2 weak cirri on the left margin. In addition, there may be a lateral ciliary row visible on either side of the ventral surface. Large fronto-ventral and transverse cirri present. Large (right) caudal cirri (usually 2-3) present. Macronucleus C-, M- or horseshoe-shaped, with single adjacent micronucleus. First described by Ehrenberg (1831). Many species have been described.
Quote; Colin R. Curds, Michael A. Gates and David McL. Roberts "British and other freshwater ciliated protozoa Part II Ciliophora: Oligohymenophora and Polyhymenophora" Cambridge University Press, 1983 (ref. ID; 2014) [ref. ID; 4401]
In Euplotes, results of mating and breeding analyses suggested the possibility of sibling species in two different circumstances: in one, to separate the two types of populations (i.e. "autogamous" and "non-autogamous", the former being capable of performing autogamy in addition to mating and, the latter being capable only of mating) which coexist in some species, such as E. minuta, E. crassus, and E. woodruffi; in the other, to partition the gene pool of a set of E. vannus strains representing only a population of the non-autogamous type. Subsequently, autogamous and non-autogamous strains in E. crassus were shown to differ in protein electrophoretic patterns and in ecological requirements as well. Mating and breeding analyses are obviously the foundation of the taxonomic procedures for identifying genetically separated populations within ciliate morphospecies. However, the results presented here suggest that the information produced by these analyses should be regarded as having a rather different weight in relation to the model of mating type system evolved, and breeding strategy pursued by the species studied. (ref. ID; 4401) [ref. ID; 4530]
In the taxonomy of the genus Euplotes, one of the main morphological traits used for carrying out a preliminary species distinction is the type of argyrome shown by the cell on its dorsal surface. This term was used by Tuffrau (1960) to indicate the geometrical, meshwork-like structure which becomes apparent after the cell's silver-staining, and reflects the lines of junction between the flat, polygonal alveoli which are joined to each other into a single layer underlying the cell plasma membrane. The diagnostic importance of the argyrome was particularly emphasized by Curds (1975) in his guide to Euplotes taxonomy. The first division of Euplotes species into groups is, in fact, made on the basis of three basic argyrome types, each defined by the number of rows of polygons lying between any two adjacent kineties. Since in different species there may be one, two, or more than two of these rows, argyromes are classified into single, double, or multiple types. Subsequently, the double argyrome type is separated into eurystomus and patella types (which are typically shown by E. eurystomus and E. patella), according to whether polygons of adjacent rows are of a similar or a different width. Based on evidence primarily derived from a morphological study of E. bisulcatus, an occasional marine species described only by Kahl (1932) and Borror (1986). (ref. ID; 4530) [ref. ID; 4932]
Differences in basic cell body traits among species of Euplotes, such as the number and disposition of the ventral cirri and the geometry of the dorsal argyrome, have been regarded as being sufficiently clear and stable to justify splitting this taxon into four morphologically distinct genera, one designated with the old name of Euplotes and the other three re-named Euplotoides, Euplotopsis and Moneuplotes (Borror and Hill 1995). (ref. ID; 4932) [ref. ID; 7309]
Morphological effects of salinity. Euryhalinity has implications for ciliate systematics. Within the genus Euplotes, most species descriptions simply report each new strain as being either marine or freshwater, with no attempt to acclimate the cells to different slinities (Curds 1975). There are several strikingly similar species discriptions differing only in their reported haitats, such as E. crenosus Tuffrau, 1960 (freshwater) and E. octocirratus Agamaliev, 1967 (marine). (ref. ID; 7309) [ref. ID; 7354]
Members of the genus Euplotes are primarily marine, however one occurs in freshwater and one occurs in the digestive tract of sea urchins. Members of this genus typically have 10 FVC and a double to complex dorsal argyrome system. The dorsal surface usually has several low dorsal ribs. There are no reported bacterial or algal endosymbionts except for a report of endosymbionts in E. moebiusi (Schlegel et al., 1988). Some members of the genus encyst. The type species is Euplotes charon (Muller, 1773) Ehrenberg, 1830. The genus Euplotes contain 25 additional species: E. alatus Kahl, 1932, E. antarcticus Fenchel & Lee, 1972, E. balteatus (Dujardin, 1841) Kahl, 1932, E. corsica Berger & Foissner, 1989, E. crenosus Tuffrau, 1960, E. euryhalinus Valbonesi and Luporini, 1990, E. focardii Valbonesi and Luporini, 1990, E. harpa Stein, 1859, E. ilifferi Hill, Small & Iliffe, 1986, E. indentatus Carter, 1972, E. inkystans Chatton in Tuffrau, 1960, E. kurekchayi Aliev, 1987, E. magnicirratus Charter, 1972, E. moebiusi Kahl, 1932. E. neapolitanus Wichterman, 1964, E. nobilii Valbonesi and Luporini, 1990, E. ogusi Alieve, 1987, E. octocirratus Agamaliev, 1957, E. platystoma Dragesco & Dragesco-Kerneis, 1986, E. polycarinatus Carter, 1972, E. quinquecarinatus Gelei, 1950, E. rariseta Curds, West, and Dorahy, 1974, E. sharuri Aliev, 1987, and E. trisulcatus Kahl, 1932. (ref. ID; 7354)
Type species; Euplotes charon (Muller, 1773) Ehrenberg, 1830 (ref. ID; 7354) [ref. ID; 7513]
Freshwater Euplotes (cirral patterns of cirrotype 9): Since Muller first described the organism now known as E. patella (Muller, 1773) Ehrenberg, 1838, there have been described at least 20 very similar species, plus at least 15 other cirrotype 9 species (Curd 1975). The advent of silver-staining (Chatton and Lwoff 1930), which reveals the infraciliature including the exact positions of the ciliary bases, created the potential for a more rigorous basis to the taxonomy of the genus. Tuffrau (1960) was the first worker to systematically examine the silver-line systems of representative samples of the genus. On the basis of his investigation of 16 species, including 3 new ones, he proposed four taxonomic criteria of paramount value in species identification. Two of these were traditional characters: the number and position of the frontoventral cirri, and the shape of the (interphase) macronucleus. To these Tuffrau (1960) added the number of dorsolateral kineties (the corticotype) and the pattern of the dorsal argentophilic network of polygonal lines (which I shall call the dargyrome). Although subsequent workes have demonstrated the variability, even among subclones, both of corticotypes (Carter 1972; Frankel 1975; Curds 1975) and of dargyromes (Gates and Curds 1978), the former two characteristics are still regarded as invariant within species of the genus (Carter 1972; Curds 1975; Hill and Reilly 1976). Both of these characteristics, cirral patterns and macronuclear shape, are usually assessed qualitatively, and this has been the source of considerable misinterpretation. Elsewhere (Gates 1976, 1977) I have demonstreated that cirral patterns the genus Euplotes can be assessed quantitatively, and that such an assessment is of value in the study of cirrotype 10 Euplotes (Gates 1978) and of the evolution of cirral pattern within the genus (Gates 1977). Washburn and Borror (1972) speculated that there are two basic cirrotype 9 patterns, depending on the location of the second cirrus in streak IV, using a modified Wallengren (1900) system of cirral enumeration. Although the positions of the other frontoventral cirri are also important in the formation of cirral configurations, this suggestion is generally confirmed when cirral patterns are analysed quantitatively (Gates 1976, 1977). In one pattern, exemplified by E. patella, there is a cirrus in the position homologous to position 2/IV in certain Euplotes samples of cirrotype 10, and no cirrus in the 3/IV position; in the second pattern, for which E. affinis provides an example, the reverse obtains. Thus, the fourth cirral streak is "missing" a cirrus in Euplotes of cirrotype 9. In fact, it is possible to demonstrate that such cirral patterns can be "derived" from certain cirrotype 10 patterns by the removal of the appropriate cirrus (Gates 1976, 1977). It is, however, only the demonstration of the internal integrity of each of these two basic cirral patterns that permits such as interpretation. (ref. ID; 7513)
- Euplotes aberrans Dragesco, 1960 (ref. ID; 2237, 3690 original paper)
See; Euplotopsis aberrans (ref. ID; 7354) - Euplotes acanthodus Petz, Song & Wilbert, 1995 (ref. ID; 4529)
- Euplotes aediculatus (Pierson, 1943) (ref. ID; 502, 1308, 1602, 1629, 2237, 2573, 3979, 4165, 4336, 4404, 4609, 4774, 7513) reported year? (ref. ID; 1618, 3763, 3995, 7750) reported author and year? (ref. ID; 191, 3292, 4471, 7400, 7688)
See; Euplotoides aediculatus (ref. ID; 7354)
Euplotes affinis-complex
[ref. ID; 7513]
Curds (1975) lists the freshwater form E. affinis and the marine form E. bisulcatus Kahl, 1932 as possessing double-eurystomus dargyrome, while two marine forms, E. apsheronicus Agamaliev, 1966 and E. zenkewitchi Burkovsky, 1970, have a double-patella dargyrome. E. muscicola Kahl, 1932, a freshwater species, is considered to have a multiple dargyrome, while E. gracilis Kahl, 1932, and E. elegans Kahl, 1932 all have complex dargytomes. The last species is euryhaline, while the previous two are freshwater forms. The multiple dargyrome of E. muscicola, in my opinion, differs from that of E. gracilis and E. muscorum Gragesco, 1970 only in being somewhat more regular. Further, the dargyrome of Tuffrau''s (1960) description of E. elegans differs from the redescription of Carter (1972); the latter is less finely divided than the former, and it conforms with the description of the above three species more closely than to the later species. (ref. ID; 7513) - Euplotes affinis (ref. ID; 191, 7309)
- Euplotes affinis Dujardin, 1841 (ref. ID; 7354) or 1842 (ref. ID; 1621, 1629, 1896, 2245, 3593) reported year? (ref. ID; 1219, 3342, 3698, 5462) or (Dujardin, 1841) Kahl, 1932 (ref. ID; 4609, 7513)
See; Euplotopsis affinis (ref. ID; 7354)
Syn; Euplotes affinis f. tricirrata Kahl, 1931 (ref. ID; 3593); Euplotes charon (O.F. Muller) Stein (ref. ID; 2245); Ploesconia affinis Dujardin, 1841 (ref. ID; 4609) - Euplotes affinis Kahl, 1932 (ref. ID; 2237)
- Euplotes affinis f. tricirrata Kahl, 1931 (ref. ID; 3593) or 1932 (ref. ID; 2237)
See; Euplotes affinis (ref. ID; 3593) - Euplotes affinis f. tricirratus Kahl, 1932 (ref. ID; 1219) reported author and year? (ref. ID; 3292)
Syn; Euplotes charon (O.F. Muller) Stein (ref. ID; 2245)
- Euplotes agamalievi (Aliev, 1987)
See; Euplotoides agamalievi (ref. ID; 7354) - Euplotes alatus Kahl, 1932 (ref. ID; 1621, 2237, 4488, 4529)
- Euplotes albus Fernandez-Leborans, 1982 (ref. ID; 4695) reported author and year? (ref. ID; 191)
- Euplotes algivora Agatha et al., 1990 (ref. ID; 4529)
- Euplotes algivorus Agatha et al., 1993 (ref. ID; 4905)
- Euplotes amieti (Dragesco, 1970) (ref. ID; 7354, 7513)
See; Euplotoides amieti (ref. ID; 7354) - Euplotes antarticus Fenchel & Lee, 1972 (ref. ID; 4529)
- Euplotes appendiculatus Ehrenberg, 1838
See; Euplotes charon (ref. ID; 2237) - Euplotes apsheronicus Agamaliev, 1966 (ref. ID; 7513)
- Euplotes balteatus (Dujardin, 1842) (ref. ID; 1621, 2237)
- Euplotes balteatus Kahl, 1932 (ref. ID; 2289, 4529)
- Euplotes balteatus (Dujardin, 1841) Kahl, 1932 (ref. ID; 7354)
- Euplotes balticus Kahl, 1932 (ref. ID; 3119) or (Kahl, 1932) Dragesco, 1966 (ref. ID; 4712)
- Euplotes bisulcatus Kahl, 1932 (ref. ID; 1621, 4530, 7513) reported year? (ref. ID; 3846)
See; Euplotes elegans (ref. ID; 2237), Euplotopsis bisulcatus (ref. ID; 7354) - Euplotes carinatus Stokes, 1885 (ref. ID; 1621, 2237) reported year? (ref. ID; 1618)
- Euplotes caudatus Meunier, 1907 (ref. ID; 1621)
See; Euplotes vannus (ref. ID; 2237) - Euplotes charon (ref. ID; 191, 7309)
- Euplotes charon Ehrenberg, 1830 (ref. ID; 2289, 4529, 4774)
- Euplotes charon (O.F. Muller, 1786) (ref. ID; 4893, 7318) reported year? (ref. ID; 1618) or (O.F. Muller, 1786) Stein (ref. ID; 1621, 2117, 2237) reported year? (ref. ID; 1219)
See; Euplotes affinis (ref. ID; 2245)
Syn; Euplotes affinis (ref. ID; 1219); Euplotes appendiculatus Ehrenberg, 1838 (ref. ID; 2237); Euplotes moebiusi Kahl (ref. ID; 2237); Trichoda cimex O.F. Muller, 1773 (ref. ID; 2237) - Euplotes corsica Berger & Foissner, 1989 (ref. ID; 4529)
- Euplotes crasus (Dujardin, 1842) (ref. ID; 1621, 1879)
Syn; Euplotes violaceus Kahl, 1928 (ref. ID; 1621) - Euplotes crassus (ref. ID; 191, 3349, 3546, 3853, 4401, 4471, 7276, 7309, 7310)
- Euplotes crassus Dujardin, 1841 (ref. ID; 2237, 3669) or 1842 (ref. ID; 3498), (Dujardin, 1841) Kahl, 1932 (ref. ID; 4712, 7513) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 7672)
- Euplotes crassus Kahl, 1932 (ref. ID; 4774)
Syn; Euplotes crassus var. minor Kahl, 1932 (ref. ID; 2237); Euplotes vannus var. balticus Kahl, 1932 (ref. ID; 2237) - Euplotes crassus var. minor (ref. ID; 3498)
- Euplotes crenosus Tuffrau, 1960 (ref. ID; 2237 original paper, 4529)
- Euplotes cristatus (ref. ID; 1621, 2237)
- Euplotes daidaleos Diller & Kounaris, 1966 (ref. ID; 4077, 4237, 4774, 7309, 7513) reported author and year? (ref. ID; 191, 7400)
See; Euplotoides daidaleos (ref. ID; 7354) - Euplotes dogieli Agamaliev, 1967 (ref. ID; 7354, 7513)
See; Euplotoides dogieli (ref. ID; 7354) - Euplotes elegans Kahl, 1932 (ref. ID; 1621, 2237, 7513)
See; Euplotopsis elegans (ref. ID; 7354)
Syn; Euplotes bisulcatus Kahl, 1932 (ref. ID; 2237) - Euplotes elegans f. litoralis (ref. ID; 1621, 2237), var. litoralis Kahl, 1932 (ref. ID; 4905)
- Euplotes elegans f. littoralis (ref. ID; 3690)
- Euplotes encysticus Yonezawa, 1985 (ref. ID; 3350) reported author and year? (ref. ID; 191)
See; Euplotopsis encysticus (ref. ID; 7354) - Euplotes euryhalinus Valbonesi & Luporini, 1990 (ref. ID; 4529)
- Euplotes eurystomus (ref. ID; 191, 3292, 7611)
- Euplotes eurystomus Kahl, 1932 (ref. ID; 4404, 4774)
- Euplotes eurystomus (Pierson) (ref. ID; 1308, 2573)
- Euplotes eurystomus Wrzesniowski, 1870 (ref. ID; 1621, 2237, 4609) reported year? (ref. ID; 1618, 5462) or (Wrzesniowski, 1870) Kahl, 1932 (ref. ID; 3119, 7513)
See; Euplotoides eurystomus (ref. ID; 7354)
Syn; Euplotes patella var. eurystomus Wrzesniowski, 1870 (ref. ID; 4609) - Euplotes eurystomus f. plumipes (ref. ID; 3292)
- Euplotes eurystomus var. marinus Agamaliev, 1971
See; Euplotes charon (ref. ID; 4893) - Euplotes excavatus Claparede & Lachmann, 1858
See; Diophrys appendiculata (ref. ID; 1621) - Euplotes extensus Fresenius, 1865 (ref. ID; 1621, 2237)
- Euplotes finki Foissner, 1982 (ref. ID; 662 original paper, 4774) reported author and year? (ref. ID; 191)
See; Euplotopsis finki (ref. ID; 7354) - Euplotes focardii Valbonesi & Luporini, 1990 (ref. ID; 4529, 4859, 6975)
- Euplotes gracilis Kahl, 1932 (ref. ID; 1621, 2237, 4404, 7513)
See; Euplotopsis gracilis (ref. ID; 7354) - Euplotes harpa Stein, 1859 (ref. ID; 1335, 1621, 2117, 2237, 3119, 4404, 4529) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 191, 3292, 7309)
Syn; Ploesconia cithara Dujardin, 1842(?) (ref. ID; 1621) - Euplotes iliffei Hill, Small & Iliffe, 1986 (ref. ID; 7354)
- Euplotes indentatus Carter, 1972 (ref. ID; 2289)
- Euplotes inkystans Chatton (ref. ID; 2237, 4529) or Chatton in Tuffrau, 1960 (ref. ID; 7309, 7354)
- Euplotes kurekchayi Aliev, 1987 (ref. ID; 4529)
- Euplotes leticiensis Bovee, 1957 (ref. ID; 1618)
See; Euplotes patella (ref. ID; 2237) - Euplotes latus Agamaliev, 1967 (ref. ID; 7513)
- Euplotes longipes Claparede & Lachmann, 1858 (ref. ID; 7354) reported year? (ref. ID; 2237)
- Euplotes magnicirratus Carter, 1972 (ref. ID; 645, 2289, 4529, 7309) reported author and year? (ref. ID; 4994)
- Euplotes margherensis Coppellotti & Cisotto, 1996 (ref. ID; 4529)
- Euplotes marioni Gourret & Roeser, 1886 (ref. ID; 1621)
See; Euplotes vannus (ref. ID; 2237) - Euplotes mediterraneus Fernandez-Leborans & Castro De Zaldumibide, 1985 (ref. ID; 4529) reported author and year? (ref. ID; 191)
- Euplotes michaellae Chardez, 1982 (ref. ID; 3561 original paper)
- Euplotes minuta Yocum, 1930 (ref. ID; 2289, 3669, 3979, 4774, 4905) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 191, 7309, 7310)
Syn; Euplotes balticus sensu Dragesco, 1966 (ref. ID; 4905); Euplotes cristatus sensu Carter, 1972 (ref. ID; 4905) - Euplotes moebiusi Kahl, 1932 (ref. ID; 3119, 4609) reported year? (ref. ID; 646, 1621, 1629) reported author and year? (ref. ID; 191)
See; Euplotes charon (ref. ID; 2237) - Euplotes moebiusi f. quadricirratus Kahl (ref. ID; 1621, 2237)
- Euplotes modunensis Balbiani, 1860 (ref. ID; 2237)
- Euplotes muscicola Kahl, 1932 (ref. ID; 662, 1621, 2237, 3593, 4195, 4488, 4861, 7513) reported author and year? (ref. ID; 7617)
See; Euplotopsis muscicola (ref. ID; 7354) - Euplotes muscicola f. alatus Kahl (ref. ID; 2237)
- Euplotes muscicola f. bialatus Kahl (ref. ID; 2237)
- Euplotes mutabilis Tuffrau, 1960 (ref. ID; 2237 original paper, 4712, 7309, 7513) reported year? (ref. ID; 3830)
See; Euplotes vannus (O.F. Muller, 1786) (ref. ID; 4893) - Euplotes neapolitanus Wichterman, 1964 (ref. ID; 4529) reported author and year? (ref. ID; 4195)
- Euplotes nobilii Valbonesi & Luporini, 1990 (ref. ID; 4529, 4905)
- Euplotes novemcarinatus Wang, 1930 (ref. ID; 1621, 2237)
See; Euplotopsis novemcarinata (ref. ID; 7354) - Euplotes octocarinatus Carter, 1972 (ref. ID; 4077, 4774) reported author and year? (ref. ID; 191, 3574, 3641, 3642, 3662, 4471, 7400, 7654)
See; Euplotoides octocarinatus (ref. ID; 7354) - Euplotes octocirratus Agamaliev, 1957 (ref. ID; 7354) or 1967 (ref. ID; 2289, 4529, 7309)
- Euplotes ogusi Aliev, 1987 (ref. ID; 4529)
- Euplotes palustris Ten Hangen, 1980 (ref. ID; 479 original paper, 4529)
See; Euplotoides palustris (ref. ID; 7354) - Euplotes paradoxa Kent, 1880
See; Euplotes patella (ref. ID; 2237) - Euplotes parawoodruffi Song & Bradbury, 1997 (ref. ID; 4518 original paper)
Syn; Euplotes woodruffi sensu Dragesco & Dragesco-Kerneis, 1986; Hill & Reill, 1976 (in part?); Magagnini & Nobili, 1964 (in part?); Borror, 1963 (?); Ikeda, 1955 (?) (ref. ID; 4518) - Euplotes parkei (ref. ID; 7309)
Euplotes patella-complex
[ref. ID; 7513]
Curds (1975) lists 5 species with the patella subtype of cirral pattern (i.e., cirrus 3/IV is missing) and a double-eurystomus dargyrome: E. aediculatus Pierson, 1943; E. amieti Dragesco, 1970; and E. eurystomus (Wrzesniowski,1870) Kahl, 1932, are the freshwater species, while E. dogieli Agamaliev, 1967 and E. latus Agamaliev, 1967 are the marine forms. On contrast, there are only 3 species, all freshwater, having the double-patella dargyrome: E. daidaleos, E. patella, and E. octocarinatus Carter, 1972 (ref. ID; 7513) - Euplotes patella syngen 1 (ref. ID; 4214)
- Euplotes patella syngen 2 (ref. ID; 4214)
- Euplotes patella (ref. ID; 191, 4750, 7400)
- Euplotes patella Ehrenberg, 1838 (ref. ID; 4077, 4404, 4774) reported year? (ref. ID; 3698, 3771, 5624), (O.F. Muller) (ref. ID; 1618), (O.F. Muller, 1773) Ehrenberg, 1831 (ref. ID; 4488, 4609) or 1838 (ref. ID; 3116), (O.F. Muller, 1773) Ehrenberg (ref. ID; 1621, 2237, 2245) reported year? (ref. ID; 1219, 3342)
Syn; Coccudina keromina Bory, 1824 (ref. ID; 2237); Euplotes carinatus Stokes, 1885 (ref. ID; 2237); Euplotes leticiensis Bovee, 1957 (ref. ID; 2237); Euplotes paradoxa Kent, 1880 (ref. ID; 2237); Euplotes patella var. lemani Dragesco, 1960 (ref. ID; 2237); Euplotes viridis Ehrenberg, 1838 (ref. ID; 2237); Trichoda patella O.F. Muller, 1773 (ref. ID; 4609) - Euplotes patella (Pierson) (ref. ID; 1308)
- Euplotes patella f. alatus Kahl, 1932 (ref. ID; 3116) reported year? (ref. ID; 1621, 2237)
- Euplotes patella f. latus Kahl, 1932 (ref. ID; 3116) reported year? (ref. ID; 1621, 2237)
- Euplotes patella f. planctonicus Kahl (ref. ID; 1621, 2237)
- Euplotes patella f. typicus Kahl, 1932 (ref. ID; 3116) reported year? (ref. ID; 1621, 2237)
- Euplotes patella f. variabilis Stokes (ref. ID; 1621)
- Euplotes patella var. eurystomus Wrzesniowski, 1870
See; Euplotes eurystomus (ref. ID; 4609) - Euplotes patella var. lemani Dragesco, 1960 (ref. ID; 3690 original paper)
See; Euplotes patella (ref. ID; 2237)
- Euplotes platystoma Dragesco & Dragesco-Kerneis, 1986 (ref. ID; 4529)
- Euplotes plicatum Valbonesi, Apone & Luporini, 1997 (ref. ID; 4529 original paper)
- Euplotes plumipes Stokes, 1884 (ref. ID; 1621, 2237, 7513) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 191, 7400)
See; Euplotoides plumipes (ref. ID; 7354) - Euplotes polycarinatus Carter, 1972 (ref. ID; 4529)
- Euplotes quinquecarinatus Gelei, 1950 (ref. ID; 2289, 7354)
- Euplotes quinquicarinatus Gelei (ref. ID; 3846)
- Euplotes raikovi Agamaliev, 1960 (ref. ID; 4774) or 1966 (ref. ID; 2289, 4893, 4905, 7354) reported author and year? (ref. ID; 191, 7111, 7730)
See; Euplotes rariseta (ref. ID; 4893), Euplotopsis raikovi (ref. ID; 7354) - Euplotes rariseta Curds, West & Dorahy, 1974 (ref. ID; 645, 2289 original paper, 4529, 4774, 4893) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 191)
Syn; Euplotes raikovi Agamaliev, 1966 (ref. ID; 4893); Euplotes streikovi Agamaliev, 1967 (ref. ID; 4893) - Euplotes roscoffensis Dragesco, 1966 (ref. ID; 2289, 7354)
See; Euplotes vannus (O.F. Muller, 1786) (ref. ID; 4893) - Euplotes shanghaiensis Song, Hill & Warren (ref. ID; 4529)
- Euplotes sharuri Aliev, 1986 (ref. ID; 4893) or 1987 (ref. ID; 7354)
See; Euplotes vannus (O.F. Muller, 1786) (ref. ID; 4893) - Euplotes strelkovi Agamaliev, 1967 (ref. ID; 2289)
See; Euplotes rariseta (ref. ID; 4893), Euplotopsis strelkovi (ref. ID; 7354) - Euplotes taylori Garnjobst, 1928 (ref. ID; 1621, 2237, 4488, 7354) reported year? (ref. ID; 4111)
- Euplotes tegulatus Tuffrau, 1960 (ref. ID; 2237 original paper, 7513)
See; Euplotopsis tegulatus (ref. ID; 7354) - Euplotes terricola Penard, 1922 (ref. ID; 1621, 2237, 7354)
- Euplotes thononensis Dragesco, 1960 (ref. ID; 3690 original paper, 7354) reported year? (ref. ID; 2237)
- Euplotes trisulcatus Kahl, 1932 (ref. ID; 1621, 1879, 2237, 3119, 4529, 7309)
- Euplotes truncatus Ehrenberg (ref. ID; 2237)
- Euplotes truncatus Meunier, 1907 (ref. ID; 1621)
- Euplotes tuffraui Berger, 1965 (ref. ID; 2289, 4529, 7354) reported year? (ref. ID; 3830)
- Euplotes turritus Ehrenberg, 1838
See; Aspidisca turrita (ref. ID; 4609)
Euplotes vannus-complex
[ref. ID; 7680]
E. crassus, E. mutabilis, and E. vannus are true sibling species? (ref. ID; 7680) - Euplotes vannus (ref. ID; 191, 902, 3629, 4471, 7309, 7310, 7509, 7525)
- Euplotes vannus Minkjewicz, 1901 (ref. ID; 2289, 4774)
- Euplotes vannus (O.F. Muller, 1786) (ref. ID; 1621, 2237, 3669, 3936, 4281, 4893) or (O.F. Muller, 1786) Minkjewicz, 1901 (ref. ID; 4712) reported year? (ref. ID; 3830)
Syn; Euplotes caudatus Meunier, 1907 (ref. ID; 2237); Euplotes crassus sensu Tuffrau, 1960 (ref. ID; 4893); Euplotes longipes Claparede & Lachmann, 1859 (ref. ID; 1621); Euplotes marioni Gourret & Roeser, 1886 (ref. ID; 2237); Euplotes minuta sensu Agamaliev, 1971 (ref. ID; 4893); Euplotes mutabilis Tuffrau, 1960 (ref. ID; 4893); Euplotes roscoffensis Drgesco, 1966 (ref. ID; 4893); Euplotes sharuri Aliev, 1986 (ref. ID; 4893); Euplotes worcesteri Griffin, 1910 (ref. ID; 1621) - Euplotes vannus var. balticus Kahl, 1932
See; Euplotes crassus (ref. ID; 2237) - Euplotes vanuus (O.F. Muller, 1786) (ref. ID; 2117)
- Euplotes variabilis Stokes (ref. ID; 2237)
- Euplotes violaceus Kahl (ref. ID; 2237)
- Euplotes viridis Ehrenberg (ref. ID; 2237)
- Euplotes woodruffi (ref. ID; 191)
- Euplotes woodruffi Gaw, 1939 (ref. ID; 2237, 4404, 4518 redescribed paper, 4774) reported year? (ref. ID; 1618)
See; Euplotes parawoodruffi (ref. ID; 4518), Euplotoides woodruffi (ref. ID; 7354) - Euplotes woodruffi (Pierson) (ref. ID; 1308)
- Euplotes worcesteri Griffin (ref. ID; 2237) reported author and year? (ref. ID; 191)
- Euplotes zenkewitchi (Burkovsky, 1970) (ref. ID; 7354, 7513) reported author and year? (ref. ID; 4773)
See; Euplotopsis zenkowitchi (ref. ID; 7354)
Euplotes aediculatus (Pierson, 1943) (ref. ID; 502, 1308, 1602, 1629, 2237, 2573, 3979, 4165, 4336, 4404, 4609, 4774, 7513) reported year? (ref. ID; 1618, 3763, 3995, 7750) reported author and year? (ref. ID; 191, 3292, 4471, 7400, 7688)
See
Euplotoides aediculatus (ref. ID; 7354)Descriptions
Elliptical; 9 frontal-ventrals; aboral surface usually without ridges, but with about 6 rows of bristles; peristome narrow; peristomal plate long triangular, drawn out posteriorly; a niche midway on the right border of peristome; anal cirri often form a straight transverse line; fourth ridge between anals may reach anterior end of body; macronucleus C-shape with a flattened part in the left-anterior region; micronucleus some distance from macronucleus at anterior-left region; post-pharyngeal sac; fresh and brackish (salinity 2.30 parts of salt per 1000) water. (ref. ID; 1618)Predotor-induced phenotypic changes. (ref. ID; 7400)
Exconjugants. (ref. ID; 7688)
Measurements
Length 110-165 um; average dimensions 132 by 84 um. (ref. ID; 1618)Euplotes affinis Dujardin, 1841 (ref. ID; 7354) or 1842 (ref. ID; 1621, 1629, 1896, 2245, 3593) reported year? (ref. ID; 1219, 3342, 3698, 5462) or (Dujardin, 1841) Kahl, 1932 (ref. ID; 4609)
See
Euplotopsis affinis (ref. ID; 7354)Synonym
Euplotes affinis f. tricirrata Kahl, 1931 (ref. ID; 3593); Euplotes charon (O.F. Muller) Stein (ref. ID; 2245); Ploesconia affinis Dujardin, 1841 (ref. ID; 4609)Descriptions
9 front-ventral cirri, 5 transverse cirri, and 4 caudal cirri; dorsal side with 5-6 ribs or keels; buccal cavity without frontal plate covering the anterior part (contrary to E. patella); macronucleus C-shaped; 1 micronucleus near the anterior end of the cell; contractile vacuole on the right side near the transverse cirri. There are some taxonomic difficulties with this organism; Tuffrau (1960) stated that E. charon and E. affinis are nonspecific. However, E. affinis as understood here shows constantly 9 front-ventral cirri, while the typical E. charon has 10 (Borror 1968). As it was not possible to find specimens possessing 10 front-ventral cirri within the populations studied by the author, it was decided to regard E. affinis Dujardin as a valid species. It should be mentioned that sometimes specimens with only 3 caudal cirri occur- i.e., the so-called E. affinis f. tricirratus Kahl, 1932. (ref. ID; 1219)Measurements
Length 40-70 um. (ref. ID; 1219)60-75x30-40 um. (ref. ID; 3342)
Euplotes albus Fernandez-Leborans, 1982 (ref. ID; 4695) reported author and year? (ref. ID; 191)
Descriptions
Paroral formation: This species has a single paroral formation (PF), composed of 22-25 oblique lines of kinetosomes. It measures 8.3 um in length and has a width of 1.2 um (avg.). The anterior end of this formation lies approximately 32 um from the apical pole of the cell, and it posterior end is located beside the posterior end of the adoral membranelle area. The number of kinetosomes varies according to the row considered (from 2 in the shortest rows to 8 in the longest). The kinetosomes of each row interrelate with those of the neighbouring rows in such a way that the kinetosomes of the ensemble of the paroral formation seem to be arranged in the intersections of a grid (Fernandez-Leborans 1982). (ref. ID; 4695)Euplotes bisulcatus Kahl, 1932 (ref. ID; 1621, 4530, 7513) reported year? (ref. ID; 3846)
See
Euplotes elegans (ref. ID; 2237), Euplotopsis bisulcatus (ref. ID; 7354)Descriptions
The only description of the argyrome of E. bisulcatus has been provided by Borror (1986); it appears to be of the double-eurystomus type, as there are two interkinetal rows of elongate, uniform polygons. However, this description was based on a study of a specimens (collected from tidal marsh ponds on the New Hampshire coast) which appear to be morphologically quite different from the specimens (collected from Kieler Bay) which Kahl (1932) originally described as E. bisulcatus. The following differences are the most important: in Borror's specimens there are prominent dorsal "double-edged ridges separated by shallow grooves" and eight dorso-lateral kineties. Kahl's specimens appear to be distinguished by a body surface engraved by two deep dorsolateral grooves (as the species name emphasizes), separated from each other by a wide central region resembling a monolithic ridge with a wide and nearly flat edge, and by five kineties; of these, one runs centrally, two on the bottom of the grooves, and two marginally. We found the E. bisulcatus specimens of our collection to be morphologically very similar to Kahl's original description of E. bisulcatus, with five kineties and the two dorsal grooves. It was decided, therefore, to complete their description with an analysis of the argyrome. This argyrome did not appear to be of the double-eurystomus type, as described for E. bisulcatus by Borror (1986), but of the double-patella type, due to the presence of longitudinal rows of narrow and wide polygons. Its organization, however, did not show any correspondence with that of other described double-patella argyromes. Going from one to the other side of the dorsal cell surface, there was no orderly alternation, typical for all other double-patella argyromes, of rows with narrow and wide polygons; in addition, adjacent rows of polygons of similar size covered a wide portion of the central region. To determine the structural basis underlying this puzzling argyrome pattern, we analyzed the "true" spatial arrangement and shape of E. bisulcatus cortical alveoli in cell serial, transverse, semi-thin sections and SEM preparations. When the argyrome pattern is considered in conjugation with the dorsal profile, it becomes clear that the alveoli are of comparable width. They appear to be of different width only on planar projection, which disregards ridges and grooves. Thus a significant material basis for defining a double-patella type of argyrome is lacking. (ref. ID; 4530)Remarks
The morphological study of E. bisulcatus and other related species of Euplotes, in particular of the structural correlation between their argyrome pattern and the profile of their dorsal surface, provided direct evidence against the validity of the distinction between double-eurystomus and double-patella argyromes. This distinction was based on observations of differences in width of the argyrome polygons in silver-stained preparations. These differences are shown here not to reflect differences in the width of the cell cortical alveoli which, instead, represent substantially uniform units of construction of the Euplotes cell surface. Rather, our work shows that this distinction, as well as inter-species variations in the geometry of argyromes of the double-patella type, should to a large extent be attributed to cell cortical ridges, which may impose sharp alterations of the cell profile, and thus cause distortions of details of the cell morphology in the microscopic image, which is a planar projection. A significant examination, closely recalling that of E. bisulcatus described here, of the role that these ridges can play in the double argyrome pattern, is provided also by E. affinis sensu Foissner et al. (1991). This species is apparently polymorphic, as it may present five to seven dorsal ridges of varied shape and size. Consistence with these variations, the argyrome has been reported by Foissner et al. (1991) to shift from a double-eurystomus to a double-patella type, as cells become distinguished by more prominent ridges. The potential use of the cell cortical ridges (and/or grooves) as a reliable diagnostic tool for the morphologic identification of Euplotes species has been repeatedly recommended in particular by Borror (1986) and Foissner et al. (1991). Unfortunately, this recommendation has usually received scant attention in most descriptions, whereby the extent to which the number and shape of ridges sculpting Euplotes surface are really invariant among conspecific cells, and hence taxonomy useful, has not been defined with certainty. Like other more unusual cell cortex protrusions, such as "wings" and "spines" described in particular in E. octocarinatus (Kuhlmann & Heckmann, 1985), they might in fact undergo intra-species modification in response to cell exposure to foreign morphogenetic stimuli (Kusch 1993; Kusch & Heckmann 1992) or, as it occurs in E. focardii (our unpublished observations), to cell transition from one nutritional stage to another. On the other hand, E. bisulcatus described in detail here provides a valid example of the importance of the patterns of cell cortical folding at useful distinctive species characters in Euplotes. Our conclusion that variations in the geometry of the Euplotes argyrome of the double type are of limited or no significance in Euplotes taxonomy is not new. The invalidity of the distinction between double-patella and double-eurystomus argyromes has been earlier proposed by Gates and Curds (1979), however, in a study whose rationale, besides being quite different, becomes now also weakened by the observations described here. This study is in fact aimed primarily at defining whether, within a clone, the relative width of two adjacent rows of argyrome polygons (as judged by the position of the "interkinetal boundary") represents a stable (hence taxonomically reliable) or a variable character. Unfortunately, it was carried out only on silver-stained specimens of an unidentified Euplotes species with a double argyrome; therefore, it is now hard to discern to what extent the significant variability that was found is intrinsic to the cell cortical alveoli or, as the evidence produced here would suggest, due to variations in the folding of the dorsal cell surface. The restriction of the basic typology of Euplotes argyromes only to the single, double, and multiple models, each one derived from the association of cortical alveoli essentially uniform in size, favors a rather parsimonious view of the evolution of Euplotes cell body constriction. Two main processes presumably occurred either independent of or coincident with each other: the duplication of the longitudinal rows of alveoli and the patterned folding of the cell cortex. The net result of these changes is an increased plasticity in the cell body construction and the acquisition of a higher surface-to-volume ratio. (ref. ID; 4530)Euplotes carinatus Stokes, 1885 (ref. ID; 1621, 2237) reported year? (ref. ID; 1618)
Descriptions
In fresh water. (ref. ID; 1618)Measurements
About 70 by 50 um. (ref. ID; 1618)Euplotes charon (O.F. Muller, 1786) (ref. ID; 4893, 7318) reported year? (ref. ID; 1618) or (O.F. Muller, 1786) Stein (ref. ID; 1621, 2117, 2237) reported year? (ref. ID; 1219)
See
Euplotes affinis (ref. ID; 2245)Synonym
Euplotes affinis (ref. ID; 1219); Euplotes appendiculatus Ehrenberg, 1838 (ref. ID; 2237); Euplotes moebiusi Kahl (ref. ID; 2237); Trichoda cimex O.F. Muller, 1773 (ref. ID; 2237)Descriptions
In salt water. (ref. ID; 1618)A morphometric anaylsis of the Euplotes charon morphotype. (ref. ID; 7318)
Measurements
70-90 um long. (ref. ID; 1618)Euplotes crassus Dujardin, 1841 (ref. ID; 2237, 3669) or 1842 (ref. ID; 3498), (Dujardin, 1841) Kahl, 1932 (ref. ID; 4712, 7513) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 7672)
Synonym
Euplotes crassus var. minor Kahl, 1932 (ref. ID; 2237); Euplotes vannus var. balticus Kahl, 1932 (ref. ID; 2237)Comments
Euplotes crassus Dujardin, 1841, E. minuta Yocom, 1930, E. vannus Muller, 1786, which are common inhabitants of sand interstices of the intertidal zone. All of them possess a single-type argyrome and a cirrotype-10 pattern. In addition, they control mating by means of high-multiple mating type systems, determined by serially dominant alleles at the single Mendelian mating type (mat) locus, and comprise populations capable of performing autogamy. The separation of E. crassus, E. minuta, and E. vannus was based on the observation of morphological characteristics (such as cell body size and shape, kinety number, kinetosome number in the central kinety, and peristomial length), and it was found consistent with result of cross-breeding tests. Most of the diagnostic traits in conventional use, however, were eventually shown to be of doubtful value because they are quite variable, even among individuals of the same clone. Environmental parameters, aging, and processes of physiological reorganization may largely contribute to this clone variation. The results obtained from the study of mating interactions, isozyme patterns and multivariate morphometrics of the Euplotes strains analyzed, provide integrated evidence that E. crassus, E. minuta, and E. vannus are good species names, each connoting one separate evolutionary entity. It was shown that between non-conspecific strains there is a very restricted gene flow, little or no sharing of particular isozyme mobilities, and perceptible morphological divergence. This divergence was evidenced by numerical taxonomic methods which must be accepted with some reservations due to both the arbitrariness of techniques an interpretations, and the difficulty in achieving statistical accuracy when relatively few morphological traits and strains are available. Nevertheless, the correspondence of the "genetic" and "biochemical" species to the "typological" species identified by multivariate analysis of the conventional taxonomic traits, provided convincing evidence for the reliability of the diagnosis which classically maintains E. minuta to be the smallest, E. crassus the intermediate, and E. vannus the largest and nearly rectangular reniform shaped species. This diagnosis was first upset in Tuffrau's revision of the genus Euplotes, because E. vannus was reported to be the species with intermediate dimension. Then it was challenged by Gates and by Machelon et al. Gates argued that the three species could really be only one -possibly to be referred to by the oldest name, E. vannus- essentially on the basis of a morphometric study of cirral patterns. This study, however, was carried out on strains previously (and, perhaps, only preliminarily) classified in other laboratory and it neglected most of the conventional diagnostic traits of Euplotes. Machelon et al. studied a set of strains collected from different geographic areas and recognized two morphospecies, which were shown to be also distinct electrophoretically, a small one (ranging in length from 53.89 to 65.89 um), and a larger one (ranging from 73.11 to 91.42 um). The former was identified with the classical E. minuta and the latter with E. vannus, described as having intermediate dimensions by Tuffrau. This "E. vannus" was though to include both the conventional taxa E. vannus and E. crassus, plus E. mutabilis (a taxon earlier recognized invalid), and to consist of a complex of five "sibling species". These species were defined on the basis of the "interfertility criterion"; however, they appeared indistinguishable from each other both morphologically and enzymologically. (ref. ID; 3669)Enzymatic activity during conjugation. (ref. ID; 7672)
Euplotes daidaleos Diller & Kounaris, 1966 (ref. ID; 4077, 4237, 4774, 7309, 7513) reported author and year? (ref. ID; 191, 7400)
See
Euplotoides daidaleos (ref. ID; 7354)Descriptions
Zoochlorellae-bearing ciliates. (ref. ID; 4237)Predotor-induced phenotypic changes. (ref. ID; 7400)
Euplotes eurystomus Wrzesniowski, 1870 (ref. ID; 1621, 2237, 4609) reported year? (ref. ID; 1618, 5462) or (Wrzesniowski, 1870) Kahl, 1932 (ref. ID; 3119, 7513)
See
Euplotoides eurystomus (ref. ID; 7354)Synonym
Euplotes patella var. eurystomus Wrzesniowski, 1870 (ref. ID; 4609)Descriptions
Elongated ellipsoid; 9 frontal-ventrals; no aboral ridges, but 7 rows of bristles; peristome wide, deep; peristomal depression sigmoid; membranellae forming sigmoid curve; end of cytophaynx far to left and anterior to the fifth anal cirrus; post-pharyngeal sac; macronucleus 3-shaped; micronucleus near flattened anterior corner of macronucleus; fresh and brackish water. (ref. ID; 1618)Measurements
Length 100-195 um; average dimensions 138 by 78 um. (ref. ID; 1618)Euplotes focardii Valbonesi & Luporini, 1990 (ref. ID; 4529, 4859, 6975)
Descriptions
- Somatic infraciliature: Our observations concerning the somatic infraciliature coincide with those of Valbonesi and Luporini (1990). However, we observe slight variability in the number of frontoventral cirri (10, rarely 9) and dorsal bristles kineties (10-11, rarely 9). (ref. ID; 6975)
- Oral infraciliature: The oral infraciliature of this species consists of a parporal membrane and an adoral zone of membranelles (AZM). The paroral membrane is located at the right posterior margin of the buccal cavity and consists of 29-37 oblique rows of kinetosomes. The AZM is formed by 39-72 paramembranelles and consists of three differently structured portions: (i) The right anterior paramembranelles. The first paramembranelle has two rows of kinetosomes, while the others have three rows each. The first row of the most anterior paramembranelles is continuous and shorter than the other two, while the first row of the remaining paramembranelles in this zone is divided into two: a shorter part on the left side and a longer one decreasing in length on the right side. The length of the membranelles increases gradually in this first part. (ii) The left anterior paramembranelles are composed of three rows of kinetosomes: two longer posterior rows and one anterior row with only four kinetosomes. All paramembranelles in this part are of the same length. (iii) The left ventral paramembranelles consist of four rows of kinetosomes; the two posterior ones are longer and have the same length, the middle one is shorter, and the most anterior one has ony three kinetosomes. (ref. ID; 6975)
- Cytoskeletal elements: In the posterior part of the buccal cavity, silver impregnation reveals a thick fibrillar bundle originating from the paroral area and extending to the left posterior AZM area. Slightly argentophilic microfibrils originate from this fibrillar structure and extend into the cytoplasm. Moreover, postoral fibers descend from the end of the buccal cavity and extend into the cell. At the ventral side, fibrillar derivatives originate from each of the cirri. Lastly, fibers originating from each transverse cirrus extend to the anterior part, connecting with a dense fibrillar node located on the anterior part of cirrus 6. Thick fibrillar derivatives pass from this node to the anterior part of the cell, while other fibrillar derivatives are laterally and posteriorly oriented. On the dorsal side, bundles of subpellicular fibers belonging to the longitudinal superficial meshwork can be seen in the anterior part of the ciliate body. (ref. ID; 6975)
- The morphogenesis of cell division: The process of cell division in Euplotes focardii can be divided into 5 stages:
- Stage 1. The oral primordium appears as a single rows of kinetosomes, proliferates, and invaginates as a pocket beneath the AZM.
- Stage 2. At this stage, the paroral membrane begins to reorganize from the right posterior side of the oral primordium. The primordia of the ventral cirri appear at the ventral side as two sets of four kinetosomal streaks (Anlagen I-IV, I'-VI'). During this stage, a group of kinetosomes constituting the Anlage of cirrus 1 (Anlagen V and V') also appear at the left side of the primordia of the ventral cirri. Morover, the fibers of the fission furrow begin to be evident in the equatorial zone of the cell.
- Stage 3. At this stage, two new kinetosomal streaks (Anlagen VI and VI') appear at the right side of the other ventral Anlagen; also, another kinetosomal streak becomes visible (Anlage VII) at the left side of the AZM of the proter.
- Stage 4. This stage is characterized by differentiation of all ventral streaks in the primordia of the ventral cirri. Thus, three cirral primordia originate from Anlagen I-IV and I'-IV', while Anlagen VI and VI' give rise to two cirral primordia. Simultaneously, Anlage VII separates into two streaks of kinetosomes that will constitute the marginal cirri of the two daughter cells (Anlagen VII and VII'). In addition, a longitudinal proliferation of the dorsal kineties occurs, following a postero-anterior gradient in the proter and antero-posterior gradient in the opisthe.
- Stage 5. Once the new ventral infraciliature structures are formed, the two sets of cirri in the proter and opisthe migrate towards their respective final positions. Also, thick fibrillar bundles associated with the new ventral cirri can be seen in both the proter and the opisthe. On the dorsal side, new caudal cirri of two daughter cells develop within the last two kineties on the right lateral side of each cell. Finally both daughter cell separate. (ref. ID; 6975)
Material examined
Euplotes focardii strain TN1 were provided by Professors A. Valbonesi and P. Luporini of Camerino, Italy. (ref. ID; 6975)Euplotes leticiensis Bovee, 1957 (ref. ID; 1618)
See
Euplotes patella (ref. ID; 2237)Descriptions
Oval, with a triangular wing on left; two right caudals furcated. (ref. ID; 1618)Measurements
87-203 (155) by 67-194 (130) um. (ref. ID; 1618)Euplotes minuta Yocum, 1930 (ref. ID; 2289, 3669, 3979, 4774, 4905) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 191, 7309, 7310)
Synonym
Euplotes balticus sensu Dragesco, 1966 (ref. ID; 4905); Euplotes cristatus sensu Carter, 1972 (ref. ID; 4905)Descriptions
See Euplotes crassus. (ref. ID; 3669)In vivo about 50-70x40-55 um; outline elongate oval with anterior end more or less narrowed; left margin more convex than right; dorso-ventrally about 1:3 flattened; usually 2 inconspicuous ventral ridges, dorsal side scarcely or only slightly ridged. Buccal field medially wide, about 3/4 of body length. Cytoplasm colourless, often contains several to many bar-like crystals but few food vacuoles. Contractile vacuole not always observed, when present, constant at level of transverse cirri near right margin. Macronucleus C-shaped or questionmark-shaped, containing several large nucleoli. Micronucleus spherical, on anterior half of macronucleus. Movement slow, slightly jerking or crawling on substrate, remains stationary for long periods. Cirri fine, frontoventral, caudal and left marginal cirri 12-15 um long, transverse cirri about 20 um long. Adoral zone of membranelles resembles that of E. vannus with its proximal portion curved at about 90 degrees angle to right; base of membranelles pretty narrow, anterior end completely on dorsal surface. Paroral membrane small, beneath cytoplasmic lip. Pharyngeal fibres curved anteriorly. Constant 10 frontoventral and 5 transverse cirri forming a "normal" pattern; 2 left marginal cirri separated and alinged evenly with small caudal cirri. Leftmost dorsal kinety containing only 5-6 basal body pairs, other kineties extending over entire length of cell, evidently sparsely ciliated. Silverline system on dorsal side irregular vannus-type. (ref. ID; 4905)
Remarks
The population studied here corresponds well with the description in the original (Yocum 1930) and in the later study (Carter 1972, called Euplotes cristatus). They differ only slightly in that the China population has a irregular silverline pattern on dorsal side and relatively lower number of basal bodies within kineties. This occurs not seldom within the genus (Song & Packroff 1997), and thus the species identification seems to be fairly certain. The specimens investigated here are slightly similar to Euplotes rariseta Curds, 1974 in general appearance. E. rariseta is, however, much smaller (vs. 30-40 um in vivo) with distinctly lower number of basal bodies in dorsal kineties (9-11 vs. 5-7) and different silverline system (vs. "double-patella" type sensu Curds 1975). Different from another similar form, Euplotes cristatus Kahl, 1932, which has been redescribed all over the world under different names (E. raikovi Agamaliev, 1966, E. rariseta sensu Petz et al., 1995; E. algivorus Agatha et al., 1993; E. nobilii Valbonesi & Luporini, 1990 and very likely E. elegans var. litoralis Kahl, 1932, too), E. minuta has both a different pattern of infraciliature, e.g. arrangement of left marginal cirri (2 vs. single one or two closely together), appearance of adoral zone of membranelles and silverline system. Agamaliev (1971) mistakenly described a psammophilic form which he found from coast of the Caspian Sea as Euplotes minuta. According to all morphological data he gave (with several mistakes in the illustrations), it is doubtless that his organism was a population of Euplotes vannus in terms of cell size, body shape, silverline system, the number of membranelles and basal bodies in dorsal kineties (q.v. Song & Packroff 1997). (ref. ID; 4905)Euplotes muscicola Kahl, 1932 (ref. ID; 662, 1621, 2237, 3593, 4195, 4488, 4861, 7513) reported author and year? (ref. ID; 7617)
See
Euplotopsis muscicola (ref. ID; 7354)Descriptions
Resting cysts: Cysts may be formed singly but more often occur in groups. A single resting cysts possesses 6-7 crests traversing the full length of the cyst surface. (ref. ID; 4195)Measurements
Resting cysts: Cysts of E. muscicola average 35.2+/-3.2 um in diameter. (ref. ID; 4195)Euplotes neapolitanus Wichterman, 1964 (ref. ID; 4529) reported author and year? (ref. ID; 4195)
Descriptions
In saltwater. (ref. ID; 4195)Euplotes octocarinatus Carter, 1972 (ref. ID; 4077, 4774) reported author and year? (ref. ID; 191, 3574, 3641, 3642, 3662, 4471, 7400, 7654)
See
Euplotoides octocarinatus (ref. ID; 7354)Descriptions
Predotor-induced phenotypic changes. (ref. ID; 7400)Escape response. (ref. ID; 7654)
Euplotes palustris Ten Hangen, 1980 (ref. ID; 479 original paper, 4529)
See
Euplotoides palustris (ref. ID; 7354)Descriptions
It is characterized by the dorsal argyrome of the "double-patella type", the small cirri IV, 2 and V, 2, the localisation of the transverse cirri in two groups, a relatively big micronucleus (diameter 3-4 um) and the ability of encystment. It does not possess any endosymbionts in its cytoplasm. Fresh-water species. (ref. ID; 479)Type locality
Euplotes palustris n. sp. found in marshy areas in a little wild-life reserve in Westfalen, FRG. (ref. ID; 479)Measurements
It measures 45-55x35-45 um. (ref. ID; 479)Euplotes parawoodruffi Song & Bradbury, 1997 (ref. ID; 4518 original paper)
Synonym
Euplotes woodruffi sensu Dragesco & Dragesco-Kerneis, 1986; Hill & Reill, 1976 (in part?); Magagnini & Nobili, 1964 (in part?); Borror, 1963 (?); Ikeda, 1955 (?) (ref. ID; 4518)Diagnosis
Large marine and brackish water Euplotes with highly arched dorsum; cells in vivo about 110-220x70-130 um, AZM over 4/5 of cell length with ca. 80 membranelles; macronucleus irregularly S- or T-shaped with left arm markedly longer than right one; 9 frontoventral, 5 transverse, 2 left marginal, and 2 caudal cirri; 8-10 dorsal kineties, each of middle ones with about 30 pairs of basal bodies. Silverline system double-eurystomus type. (ref. ID; 4518)Descriptions
Cells in vivo ca. 120-160 um in length; body shape more or less triangular or (less frequently) somewhat rectangular in large specimens, possibly close to division, with anterior end slightly to conspicuously wider than posterior; posterior end asymmetrically narrowed in slim individuals; both left and right side almost straight. Anterior margin shoulder-like because of sharply angled adoral zone of membranelles, especially noticeable on anterior right where the AZM is indented slightly. Prominent plasmatic protrusion about 20 um high, collar-like and hyaline, paralleling AZM dorsally on anterior margin. Body dorsoventrally thin and flattened (especially caudal region behind transverse cirri), but arched strongly (convex dorsally and concave ventrally), semicircular in outline when viewed apically, thus resembling Euplotidium at first glance. Buccal field spacious and conspicuously long, extending approximately 80%-90% of body length; midway along right border, buccal lip indented slightly, forming a large, triangular buccal cavity. Anterior portion of buccal area hyaline with diagonally positioned "step" at about 1/2 height of buccal field. Right buccal overture with wide ventral lip, extending right from anterior-most corner of buccal field and ending to left of left-most transverse cirrus. No evident ridges on ventral side except those between transverse cirri. On dorsal surface neither ridges not grooves detectable (both in vivo or after impregnation). Numerous fine granules (ca. 1 um across) (mitochondria?) immediately beneath pellicle (like in E. woodruffi), packed together on dorsal surface in polygonal mosaic pattern, but when sparser, being oval to ellipsoid with irregular gaps among them. Cytoplasm colourless, but right of buccal field often filled with several large food vacuoles (10-20 um) and tiny granules making cytoplasm dark grey. Contractile vacuole anterior to right of transverse cirri. Locomotion by slowly crawling on substrate, occasionally remaining stationary for rather long periods. Macronucleus usually more like irregular "S" than "T", thin compared with that of E. woodruffi, containing many small nucleoli and centrally located, "vacuoles" which are only weakly stained by protargol. Upper portion of stem directed to right, two arms obviously unequal length (right arm only about 1/3-1/2 of left one). Prominent left arm directly beneath adoral zone of membranelles and terminating close to hook-shaped protrusion on stem. Micronucleus small, located usually between distal end of left arm and stem hook, in most cases only faintly impregnated by protargol. Cirri thicker than those of E. woodruffi: frontoventral cirri ca. 35 um long, transverse cirri 45-55 um, caudal and marginal cirri 30-35 um. Fimbriation of caudal cirri distinct. Dorsal kinety 1 shortened anteriad, other dorsal kineties extending over entire length of cell, containing ca. 30 kinetosome pairs in each kinety and anterior ends abutting to AZM. Adoral zone of membranelles markedly wide (bases of membranelles in main portion ca. 18+/-2 um long), anteriorly almost at right-angles on both left and right cell margins with its posterior portion abruptly narrowed. Distal end of AZM curved ventrally, into/between anterior frontal cirri; posterior end terminating in buccal cavity beneath right-most transverse cirrus. Paroral membrane similar to that of E. woodruffi, medium long, with anterior portion often twisted and comb-like. Silverline system on dorsal surface regular double-eurystomus type. Contractile vacuole pore right of transverse cirri. (ref. ID; 4518)Comparison of E. parawoodruffi and E. woodrufii
Euplotes parawoodruffi differs from all other known congeners except E. woodruffi in the unique shape of the macronucleus. (ref. ID; 4518)- Body shape: Like all freshwater strains found both in China and elsewhere, Euplotes woodruffi is oval in outline contracted to the triangular/rectangular shape of E. parawoodruffi. The very large buccal field in E. parawoodruffi (over 4/5 of body length differs from the buccal field of only about 2/3 of body length in woodruffi. Gaw's original description (Gaw 1939) and subsequent studies (Hill & Reill 1976; Hosoi 1973; Shi & Wang 1989) in addition to our observations establish that the freshwater form (E. woodruffi) has a dorso-ventrally flattened body shape with inconspicuous dorsal grooves. In E. parawoodruffi, the cell is C-shaped in cross-section like an Euplotidium and has no ridges or grooves on the dorsal surface.
- Pre-oral pouch: This critical feature, conspicuously present in E. woodruffi, was only recently described (Shi & Wang 1989). This narrow, teardrop-shaped invagination on the dorsal wall of the buccal cavity anterior to the cytostome is definitely absent in parawoodruffi.
- Adoral zone membranelles (AZM): The constant differences observed in the appearance of the AZM are the typically "7-shaped" AZM in parawoodruffi with the distal end bending sharply to the right and the proximal portion left of the buccal field on the ventral surface being "rapidly" narrowed posteriad. By contrast, the main part of the AZM on the left margin of E. woodruffi is more or less evenly curved (instead of straight) with its posterior part only gradually confined. The width of the AZM differs in two species; although the cells are about the small size, the (longest) bases of membranelles in woodruffi are considerably shorter than those in parawoodruffi (usually 13-15 vs. 18-20 um). In addition, in parawoodruffi the anterior (distal) end of the AZM inserts ventrally in between anterior frontal cirri, whereas in woodruffi it is not (or only slightly) curved to the ventral surface. Since the buccal field of parawoodruffi is relative much longer than that of woodruffi, the proximal end of AZM in parawoodruffi is usually immediately beneath the position of the left-most transverse cirrus on the surface while in woodruffi it is usually anterior to it with an intervening space.
- Macronucleus: The conspicuous dissimilarity in the length of the macronuclear arms is a major difference between the two species. In Euplotes parawoodruffi the right arm is usually half or less the length of the left arm, so the stem always connects the arms at the upper right corner of the cell, lending a S-shape to the whole macronucleus. In addition, the macronuclear arms and stem are thinner than in E. woodruffi. In woodruffi, both arms are short and about the same length, or the right arm may be (rather often) slightly longer. The stem joins the arms approximately in the midline of the body, giving the macronucleus a T-shape.
- Ciliature and silverline system: The ciliature and silverline system in both species are very similar, but some differences exist in the dorsal kineties. The kineties in E. parawoodruffi contain generally more basal body pairs than in E. woodruffi (on average 30 vs. 25); the distance between anterior end of each kinety and adoral zone of membranelles in parawoodruffi is consistently much shorter than that in woodruffi. (ref. ID; 4518)
Remarks
Dragesco & Dragesco-Kerneis reported a Euplotes (under the name of E. woodruffi) found from brackish water in Africa (1986) and described according to silver impregnations. It resembles our population in most characteristics, e.g. the pattern of ciliature, the appearance of the AZM, the shape of the macronucleus, and other morphometric features. It seems certain that this population is identical to E. parawoodruffi. In 1963, Borror described briefly a Euplotes woodruffi population from the coast of Florida. His schematized illustrations are not complete enough to link it definitely to E. parawoodruffi, but based on the marine biotope and some morphometric characteristics, it may be the same species. Two populations of Euplotes woodruffi were described by Magagnini & Nobili in 1964 ("one dwelling in sand at the mouth of the Serchio river, the other living in semi-stagnant water near Pisa"). Unfortunately, all drawing seem from shrunken silver impregnated specimens without giving further biotope information. We suggest that these two populations are two different forms equivalent to E. woodruffi sense Auct and E. parawoodruffi described here. The specimens with dorsal kineties extending close to the AZM are like E. parawoodruffi. The form with a short left arm of the macronucleus resembles a "true" E. woodruffi although the ventral side is perhaps from a cell in morphogenesis -the oral primordium is visible posterior to the buccal field- so perhaps it is a dividing E. parawoodruffi. Probably Ikeda's population (Ikeda 1955) is also conspecific with our E. parawoodruffi though it is depicted as having an oval body shape (schematized?) and no information on infraciliature or silverline system was given. It is recognizable as E. parawoodruffi by the appearance of the AZM, the macronucleus, and the biotope. (ref. ID; 4518)Type location
Occurred (3/9/1996) in Pamlico Sound near Aurora, North Carolina, USA. Salinity about 5 0/00. (ref. ID; 4518)Type specimens
One slide of holotype (with protargol stained specimens) and one paratype (slide as silver nitrate impregnated specimens) are deposited in the collection of WS. A duplicate set (as protargol and silver nitrate impregnated slides) is deposited in the protozoan type slide collections of the National Museum of National History, Smithsonian Institution, Washington. D.C. (ref. ID; 4518)Euplotes parkei (ref. ID; 7309)
Descriptions
The species is a cirrotype eight (though occasional individuals within a clonal strain possess a ninth cirrus), double dargyrome species, roughly 40 um in length and 30 um in width. (ref. ID; 7309)Type locality
Sampled from a freshwater source in Vienna, Austria. (ref. ID; 7309)Euplotes patella (ref. ID; 191, 4750, 7400)
Descriptions
Predotor-induced phenotypic changes. (ref. ID; 7400)Euplotes patella Ehrenberg, 1838 (ref. ID; 4077, 4404, 4774) reported year? (ref. ID; 3698, 3771, 5624), (O.F. Muller) (ref. ID; 1618), (O.F. Muller, 1773) Ehrenberg, 1831 (ref. ID; 4488, 4609) or 1838 (ref. ID; 3116), (O.F. Muller, 1773) Ehrenberg (ref. ID; 1621, 2237, 2245) reported year? (ref. ID; 1219, 3342)
Synonym
Coccudina keromina Bory, 1824 (ref. ID; 2237); Euplotes carinatus Stokes, 1885 (ref. ID; 2237); Euplotes leticiensis Bovee, 1957 (ref. ID; 2237); Euplotes paradoxa Kent, 1880 (ref. ID; 2237); Euplotes patella var. lemani Dragesco, 1960 (ref. ID; 2237); Euplotes viridis Ehrenberg, 1838 (ref. ID; 2237); Trichoda patella O.F. Muller, 1773 (ref. ID; 4609)Descriptions
Ventral side shows characteristic set of cirri somatic cirri (front-ventral: 9, transversal: 5, caudal: 4) and buccal cirri; the dorsal side bears small sensory bristles the base of which is marked by small granules forming asterisks, otherwise the dorsal side is bare but very often exhibits 5-7 ribs of keels of varying height, the height of the ribs as well as the contour of the whole body varies greatly; symbiotic zoochlorellae are sometimes present; anterior part of the buccal area covered by a rigid plate; macronucleus is C-shaped; 1 small micronucleus in the anterior part of the cell above the buccal cavity; 1 contractile vacuole on the right side near the transverse cirri. (ref. ID; 1219)Subcircular to elliptical; 9 frontral-ventrals; aboral surface with 6 prominent ridges with rows of bristles embedded in rosettes of granules; peristome narrow; peristomal plate small triangle; macronucleus simple C-form band; micronucleus near anterior-left end; membranellae straight; posterior end of cytopharynx anterior to, and to left of, the fifth anal cirrus; post-pharyngeal sac; fresh and brackish water. (ref. ID; 1618)
Measurements
Length 80-150 um. (ref. ID; 1219)Average dimensions 91 by 52 um. (ref. ID; 1618)
100-110x47 um. (ref. ID; 3342)
Euplotes plicatum Valbonesi, Apone & Luporini, 1997 (ref. ID; 4529 original paper)
Diagnosis
Marine, small-size Euplotes. Cell dorsal surface with a pleated profile due to 7 (rarely, 8) prominent ridges sharply bent distally. Ten fronto-ventral, 5 transverse, and 4 caudal cirri. Adoral zone with 22-25 ciliary membranelles. Ten kineties and an argyrome of double type, with uniform rows of polygons. One hook-shaped macronucleus and one spherical micronucleus. (ref. ID; 4529)Descriptions
The cell body is oval shaped, with the left margin more convex than the right one. The dimensions vary from 42 to 55 um in length and from 24 to 40 um in width. The cell dorsal surface has a pleated aspect due to 7 broad and prominent ridges showing their distal portions a sharply bent towards the left side of the cell. One additional ridge, making a total of 8, located on the right half of the cell surface, has been observed in a small percentage of specimens. Close to left side of each one of these ridges there is one kinety formed by ciliferous kinetosomes, usually amounting to 14 in the central (or 5th) kinety. Three other kineties, one dorsal close to the cell right margin and 2 ventro-lateral, align non-ciliferous kinetosomes. The argyrome is of double type, with elongated polygons (that reflect the boundaries of the alveolar sacs underlying the plasma membrane) similar in size and shape. These polygons amount to 15 in the two centralmost interkinetal rows. The cell ventral surface is stretched by 4 extended (or complete) and 2 shorter (or incomplete) rides. It bears a peristomial field that occupies about two-thirds of the body length and is surrounded by 22-25 adoral membranelles. The first complete ridge (counting from the left side of the body) is the broadest one. It runs adjacent to the inner border of the peristomial field and terminates posteriorly on the left of cirrus 1/I. The second and the third ones separate cirri of streaks I and II, and of steaks II and III, respectively. The fourth one lies between cirri 2 and 3 of streak IV and terminates posteriorly on the right side of cirrus 1/V. The two incomplete ridges are confined to the posterior end of the cell ventral surface, where they emerge at either side of cirrus I/IV. The cirral pattern consists of 10 fronto-ventral, 5 transverse, and 4 caudal cirri, two of which emerge from the rear of the cell, and two are shifted towards the cell left side. In starved cells, arrested in the G1 (or, G0) stage of the cell cycle, the nuclear apparatus consists of one hook-shaped macronucleus and one spherical micronucleus lying close to the left anterior edge of the cell body. (ref. ID; 4529)Taxonomy
Euplotes plicatum falls into a group of 27 Euplotes species characterized by 10 fronto-ventral cirri and a dorsal argyrome of double type. (We refrain here from introducing a further distinction of these species into the double-patella and double-eurystomus subgroups, since this distinction is taxonomically unreliable, as earlier discussed, in particular by Gates & Curds 1979, and Valbonesi & Luporini 1995). The diagnosis of these species has been essentially based on the comparative analysis of the morphological traits. These traits are reliable only if they diverge sufficiently from one to another species to warrant their use, and are considered as a whole (not individually). Most of them are in fact subjected to marked variations even among individuals of the same species (Curds 1975; Hill & Reilly 1976; Gates 1978). Thus measures and qualitative attributes of each one of the 27 Euplotes species of the same group as E. plicatum have been first coded into a few states with a wide interval of variability, and then jointly analyzed by means of an "unweighted pair-group method using arithmetic averages" (UPGMA) which has been applied to a similarity matrix derived from Gower's similarity coefficients. This cluster analysis, in spite of its tendency to compress rather than to widen inter-species differences (as a matter of fact, E. alatus, E. algivora, E. corsica, E. nobilii, and E. trisulcatus, would appear to be not separate from one another), produces a dendrogram showing that E. plicatum clearly diverges from all the other species. The species closest to E. plicatum appears to be E. euryhalinus, linked at 83% of similarity. This extent of similarity would have been even lower, had E. plicatum and E. euryhalinus been compared also for the aspect of these dorsal surfaces, nearly smooth in the former versus pleated in the latter. Notwithstanding a recognized value in Euplotes taxonomy (Borror 1986; Foissner et al. 1991; Valbonesi & Luporini 1995), a description of the architecture of the cell dorsal surface is not available for every species; thus, this trait could not be included in the cluster analysis. (ref. ID; 4529)Etymology
This species denomination is derived from the latin "plicatum" (pleated); it was chosen to emphasize the peculiar pleated silhouette of the cell dorsal surface. (ref. ID; 4529)Type specimens
The holotype and a paratype of E. plicatum of silver-stained cells have been deposited at the British Museum (National History) in London (the accession number has not yet been assigned). A reference strain (NZ2) has been deposited at the Culture Collection of Algae and Protozoan (The Windermere Laboratory, Amblleside, Cumbrial LA22 OLP, UK) and culture of E. plicatum are available from our laboratory. (ref. ID; 4529)Euplotes plumipes Stokes, 1884 (ref. ID; 1621, 2237, 7513) reported year? (ref. ID; 1618) reported author and year? (ref. ID; 191, 7400)
See
Euplotoides plumipes (ref. ID; 7354)Descriptions
Similar to E. eurystomus. In fresh water. (ref. ID; 1618)Predotor-induced phenotypic changes. (ref. ID; 7400)
Remarks
E. plumipes of Tuffrau (1960), which is now considered to be a synonym of E. eurystomus (Pierson, Gierke and Fisher 1968; Curds 1975; Hill and Reilly 1976): large size; a wide, tapering peristome with a long, sigmoidal AZM and high, prominent anterior collar; an anterior peristomial pouch; a 3-shaped macronucleus; and even a classical double-eurystomus dargyrome. In contrast, the Wtaq and Jemu samples each represent a classical E. patella: medium to large size; a wide, triangular peristome, with a shorter, straighter AZM; a C-shaped macronucleus; and a double-patella dargyrome. (ref. ID; 7513)Measurements
About 125 um long. (ref. ID; 1618)Euplotes raikovi Agamaliev, 1960 (ref. ID; 4774) or 1966 (ref. ID; 2289, 4893, 4905, 7354) reported author and year? (ref. ID; 191, 7111, 7730)
See
Euplotes rariseta (ref. ID; 4893), Euplotopsis raikovi (ref. ID; 7354)Descriptions
Marine species. (ref. ID; 7111)Euplotes rariseta Curds, West & Dorahy (ref. ID; 645, 2289 original paper, 4529, 4774, 4893) reported year? (ref. ID; 3830) reported author and year? (ref. ID; 191)
Synonym
Euplotes raikovi Agamaliev, 1966 (ref. ID; 4893); Euplotes streikovi Agamaliev, 1967 (ref. ID; 4893)Diagnosis
Small (30-45 um long, 20-31 um wide), ovoid marine hypotrich with 10 fronto-ventral, 5 transversal and 3 caudal cirri; caudal cirrus below AZM stout. Ventral surface heavily sculptured with 6 posteriorly projecting ridges. Dorsal surface with 6 double-edged longitudinal ridges. Dorsal bristles sparse; 6 kineties with a maximum of 6 bristles in the central kineties. Dorsal silver-line system double of the patella type. Macronucleus S-shaped. Micronucleus small, situated anteriorly. (ref. ID; 2289)Descriptions
Only 25 to 30 um in length, E. rariseta is one of the smallest species of the genus. The dorsal surface is slightly rounded, with 4 ribs. The end of the cell is flattened both dorsally and ventrally. On the ventral surface are 10 frontoventral, 5 transverse and 3 caudal cirri. The adoral membranelle zone runs is a straight line parallel to the left edge of the animal as far as the middle of the body, and then bends sharply toward the cytostome. It consists of 20 to 25 membranelles. The macronucleus is C-shaped, with a micronucleus. The dorsal silverline system is of the double-patella type. It links 5 dorsal kineties with at most 6 cilia. The individuals described by Curds et al. (1974) have 6 dorsal kineties and one S-shaped macronucleus. (ref. ID; 645) [ref. ID; 2289]The dorsal surface is conspicuously sculptured with 6 double-edged longitudinal ridges. To the left of each ridges is a parallel row of pits from which the short (2 um long) dorsal cilia or bristles emerge. The ventral surface is also strongly sculptured, particularly in the posterior region between the transverse cirri. There are 6 ventral ridges, 3 of which are prominent and consist of an oblique midventral ridge, and 2 ridges which run longitudinally flanking the 2 outermost transverse cirri. The other 3 ridges are less prominent and are more or less restricted to the area immediately between the transverse cirri. All 6 ventral ridges project slightly posteriorly between the transverse cirri. All 6 ventral edges project slightly posteriorly between the transverse cirri and form fin-like structures. The right buccal overture has a slight anterior evagination as in Euplotes alatus Kahl, 1932 but in the species under description it extends only just past the AZM. (ref. ID; 2289)
- Ciliary organelles: This Euplotes has 10 frontoventral cirri. The arrangement closely resembles that of both Euplotes charon Ehrenberg, 1830 and Euplotes quinquecarinatus Gelei, 1950. There is the normal complement of 5 transverse cirri which arise between the ventral ridges and these, particularly cirri III (I) and IV (I), are frequently seen to be frayed at the distal end. The tendency for the transverse cirri to disrupt in this way is greatly increased during fixation. There are only 3 caudal cirri, and in this species the one below and nearest to the AZM is characteristically held stiffly out to the left side and is much stouter than the other two caudal cirri which arise beneath the central transverse cirri. The presence of this large stiff caudal cirrus appears to be a feature unique to this species; in the living cell, the cirrus seems to act as a rudder-like structure. The dorsal bristles are sparse in this species; there are 6 rows and the mid-dorsal rows have only 6 bristles, in which respect this species resembles Euplotes bisulcatus Kahl, 1932. (ref. ID; 2289)
- Nuclei: The macronucleus of this species is an irregular S-shape. In this respect it appears to be unique in the genus Euplotes. The posterior part of the macronucleus points anteriorly and ventrally towards the position of the cytostome. The micronucleus is situated close to the macronucleus on the left anterior edge of the body. (ref. ID; 2289)
- Silver-line system: The geometry of the dorsal silver-line system is of the double of patella-type and closely resembles that of Euplotes raikovi Agamaliev, 1966 with a series of alternate longitudinal rows of narrow long polygons and wide short polygons. The ventral argyrome consists of an irregular network of polygons in similar numbers to that of E. cristatus Kahl, 1932. The dorsal silver-line network has been seen on some scanning-electron micrographs. (ref. ID; 2289)
Type slides
Type slides showing silver-line systems, nuclear apparatus and whole mounts have been deposited in the slide collection of the Protozoa Section, BM (NH): holotype Reg. No. 1972:II:I:II, paratype Reg. Nos. 1972:II:I:I2-I8. (ref. ID; 2289)Measurements
Euplotes cells from the logarithmic phase are considerably larger (42+/-3 um long by 28+/-3 um wide) than are cells from the stationary phase (34.6 um long by 24.2+/-2.3 um wide). (ref. ID; 2289)Euplotes taylori Garnjobst, 1928 (ref. ID; 1621, 2237, 4488, 7354) reported year? (ref. ID; 4111)
Descriptions
Resting cyst: Empty cyst showing slit through which organism escaped, three wall layers present, outer layer wrinkled. (ref. ID; 4111)Euplotes tegulatus Tuffrau, 1960 (ref. ID; 2237 original paper, 7513)
See
Euplotopsis tegulatus (ref. ID; 7354)Remarks
Tuffrau (1960) described a distinctive, highly ridged, marine species, which appears to lack cirrus 2/III, and which has a unique dargyrome (Curds 1975). (ref. ID; 7513)Euplotes vannus (ref. ID; 191, 902, 3629, 4471, 7309, 7310, 7509, 7525)
Descriptions
The position of starlike structures ("sensory cilia") in the pellicle of Euplotes vannus in vivo. (ref. ID; 7525)Euplotes vannus (O.F. Muller, 1786) (ref. ID; 1621, 2237, 3669, 3936, 4281, 4893) or (O.F. Muller, 1786) Minkjewicz, 1901 (ref. ID; 4712) reported year? (ref. ID; 3830)
Synonym
Euplotes caudatus Meunier, 1907 (ref. ID; 2237); Euplotes crassus sensu Tuffrau, 1960 (ref. ID; 4893); Euplotes longipes Claparede & Lachmann, 1859 (ref. ID; 1621); Euplotes marioni Gourret & Roeser, 1886 (ref. ID; 2237); Euplotes minuta sensu Agamaliev, 1971 (ref. ID; 4893); Euplotes mutabilis Tuffrau, 1960 (ref. ID; 4893); Euplotes roscoffensis Drgesco, 1966 (ref. ID; 4893); Euplotes sharuri Aliev, 1986 (ref. ID; 4893); Euplotes worcesteri Griffin, 1910 (ref. ID; 1621)Descriptions
See Euplotes crassus. (ref. ID; 3669)They are marine, sand-dwelling ciliates, 80-100 um long, with 10 fronto-ventral cirri, at least eight dorsal ciliary rows, and a single argyrome. The discovery of mating types (Heckmann 1963, 1964) has provided a powerful tool for investigating the relationships among these organism of similar appearance. Since intraclonal processes are very uncommon and since the main sexual process is interclonal conjugation, the interfertility criterion can be easily applied. According to this criterion, whole strains have proved to belong to at least five intrafertile and intersterile groups making up the E. vannus complex (Genermont et al. 1976; Machelon 1978; Machelon 1982). These groups cannot be easily separated on the basis of any morphological characteristic. Thus the E. vannus complex is made up of five essentially reproductively but not necessarily geographically isolated subunits which, being morphologically similar, can be considered as sibling species. (ref. ID; 4281)
Euplotes woodruffi Gaw, 1939 (ref. ID; 2237, 4404, 4518 redescribed paper, 4774) reported year? (ref. ID; 1618)
See
Euplotes parawoodruffi (ref. ID; 4518), Euplotoides woodruffi (ref. ID; 7354)Descriptions
Oval; 9 frontal-ventrals; aboral surface often with 8 low ridges; peristome wide, with a small peristomal plate; end of cytopharynx almost below the median ridge; fourth ridge between anal cirri often extends to anterior end of body; post-pharyngeal sac; macronucleus consistently T-shaped; micronucleus anterior-right; brackish (with salinity 2.30 parts of salt per 1000) and fresh water. (ref. ID; 1618)Redescription
Improved diagnosis: Large, freshwater Euplotes in vivo 110-180x80-130 um; body shape oval, flattened dorsoventrally with inconspicuous dorsal grooves; buccal field about 2/3 of cell length with ca. 60-70 membranelles; 9 frontoventral, 5 transverse, 2 left marginal and 2 caudal cirri; about 10 dorsal kineties, each middle kinety with ca. 25 pairs of basal bodies; macronucleus T-shaped; silverline system double-eurystomus type. (ref. ID; 4518)[Morphology of live and impregnated specimens]: Our population in vivo 130-160x90-110 um in length; cells asymmetrically oval with anterior portion slightly wider than posterior; left margin usually less convex than right one, especially in well-fed cells; dorsally on anterior margin a hyaline collar-like plasmatic protrusion (about 15 um in height) at base of adoral zone of membranelles. Body dorsoventrally flattened (about 3-4:1). Buccal field wider, extending approximately 2/3 of cell length. Slight indentation in right buccal lip at anterior-most region; pre-oral pouch ("posterior dorsal recess" of Shi & Wang 1989) in shape of inverted teardrop (about 15 um in length), invaginating on dorsal wall of buccal cavity. Five short, inconspicuous ridges between transverse cirri on ventral side; 8 shallow grooves (not ridges as in earlier reports) on dorsal surface, often undetectable in wall fed individuals. Numerous ellipsoid granules (mitochondria?) ca. 1 um long, extremely densely packed and forming a mosaic pattern immediately beneath dorsal surface. Cytoplasm colourless, highly transparent, often filled with several to many different-sized lipid droplets (1-4 um dia.). Food vacuoles usually 5-30 um across, few in number, containing (depending on food resources) green algae, bacteria, or other undetermined stuffs rendering cell centre dark grey to black. Large contractile vacuole (CV) to right of anterior-most transverse cirrus. Locomotion typical of large Euplotes, usually slowly crawling on substrate, occasionally remaining stationary for short periods. Macronucleus containing numerous small nucleoli, characteristically T-shaped, with two shorter arms and slightly curved "stem"; shape variable, right arm generally longer than left; sometimes hook-shaped protrusion in middle of stem. Micronucleus between distal end of left arm and hook, faintly impregnated by protargol. Cirral pattern as described by previous authors (Hill & Reilly 1976; Hosoi 1973; Ikeda 1955; Shi & Wang 1989). Cirri thin and weak; right most-two caudal cirri about 40 um long, with fimbriated tips; left marginal and transverse cirri ca. 40-50 um long; all other fronto-ventral cirri about 40 um long. Always two "dorsal" kineties visible on ventral surface; other dorsal kineties extending over entire length of dorsal surface, each containing on average ca. 25 cilia; anterior ends of those kineties always distant from adoral zone of membranelles. Adoral zone of membranelles (AZM) evenly curved on left cell margin with its posterior portion gradually narrowed. Except at collar and posterior end, bases of membranelles about 11-15 um long; distal end of AZM curved only slightly to ventral side, thus usually not into/between anterior frontal cirri; posterior end (beneath buccal lip) terminating generally anterior or right of transverse cirri with conspicuous distance. Paroral membrane long, with anterior portion twisted and comb-like. Silverline system regular double-eurystomus type (Curds 1975) on dorsal surface. Contractile vacuole pore at right of transverse cirri. (ref. ID; 4518)