Coleps
Coleps Nitzsch, 1817 (ref. ID; 3540), 1827 (ref. ID; 2013, 4730) or 1870 (ref. ID; 3690)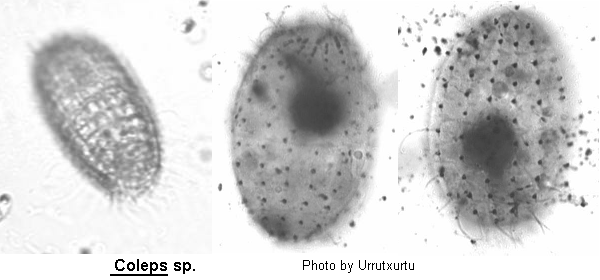
Order Prostomatida (ref. ID; 7040) [ref. ID; 2013]
Body barrel-shaped, covered in regularly arranged prominent ectoplasmic plates that are composed of amorphous calcium carbonate. Each end of the cell is rounded or slightly flattened, never pointed but often with tooth-like projections from the plates. Oral aperture circular, apically situated. Somatic ciliature uniform in regular longitudinal kineties along the striations in the plates. One or more caudal cilia present but easily over-looked. Anterior end with 2 rings of peribuccal cilia and a dorsal brush. Macronucleus ovoid, contractile vacuole posterior.
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013)
[ref. ID; 7728]
Studies on the morphology, food selection and growth. (ref. ID; 7728)
[ref. ID; 7759]
The genus Coleps are characterized by a calcified armour. Its chemical composition has been described by Faure-Fremiet et al. (1968). The armour is composed of endoskeletal plates which lie in subpellicular alveoli. The plates are arranged in six girdles. This anteriormost girdle, surrounding the cytostome, consists of regularly formed small circumoral plates, the posterior-most is formed by irregular caudal plates, some of which have spines. The plate of the four girdles between the circumoral and the caudal girdle resemble each other. They differ in size. In some species, e.g. Coleps hirtus, the secondary plates are shorter than the main plates. In addition, the plates of the anterior secondary girdle taper at their anterior part (Maupas 1885; Noland 1925; Kahl 1930). Species-sepcific differences exist in plate morphology and/or number and size of the endoskeletal plates. Based on investigations by light microscopy Kahl (1935) distinguished three different types of plate in the genus Coleps. The main characteristics are the position of the longitudinal ridge (at the right side or shifted to the middle) and the spatial arrangement of the "windows" and the lateral "teeth". Type 1 is found in freshwater species, e.g. C. hirtus, C. amphacanthus, C. elongatus, type 2 occurs only in C. nolandi and type 3 is found in the marine species C. pulcher, C. tesselatus and in the freshwater species C. incurvus. (ref. ID; 7759)
- Coleps amphacanthus Ehrenberg, 1833 (ref. ID; 1335, 1619, 2141, 3540, 4266) reported year? (ref. ID; 4613) reported author and year? (ref. ID; 191, 4611)
Syn; Coleps uncinatus Roux, 1889 (ref. ID; 1619, 3540) - Coleps bicuspis Noland, 1925 (ref. ID; 1619, 2141, 2245, 3540) reported year? (ref. ID; 1618, 4613) reported author and year? (ref. ID; 4266)
- Coleps dentatus Lepsi, 1965 (ref. ID; 2141)
- Coleps elongatus Ehrenberg, 1830 (ref. ID; 1619, 2100, 2141, 3540) or 1831 (ref. ID; 4613) reported year? (ref. ID; 1618, 3690) reported author and year? (ref. ID; 4611)
- Coleps heteracanthus Noland, 1937 (ref. ID; 2141) reported year? (ref. ID; 1618)
- Coleps hirtus O.F. Muller, 1786 (ref. ID; 4730) reported year? (ref. ID; 1335, 1618), (O.F. Muller, 1786) Nitzsch, 1827 (ref. ID; 4611) or Nitzsch, 1817 (ref. ID; 1219, 1308, 1619, 1879, 1890, 2100, 2141, 2245, 3115, 3540) reported year? (ref. ID; 3342, 3690) reported author and year? (ref. ID; 191, 5388)
Syn; Cercaria hirta Nitsch, 1817 (ref. ID; 4730); Cercaria hirta O.F. Muller, 1786 (ref. ID; 1619, 3115, 3540, 4611, 4730); Coleps incurvus Ehrenberg, 1841 (ref. ID; 3115); Coleps viridis Ehrenberg, 1838 (ref. ID; 1619, 3115, 3540); Dictiocoleps hirtus Diesign, 1866 (ref. ID; 3115); Vorticella punctata Abildgaard, 1793 (ref. ID; 3540) - Coleps hirtus hirtus (O.F. Muller, 1786) Nitzsch, 1827 (ref. ID; 4613 redescribed paper)
Syn; Cercaria hirta O.F. Muller, 1786 (ref. ID; 4613) - Coleps hirtus var. lacustris Faure-Fremiet, 1924 (ref. ID; 1619, 2141, 3540) reported author and year? (ref. ID; 3292)
- Coleps hirtus var. minor Kahl, 1930 (ref. ID; 1619, 2141, 3540 original paper, 4611, 4613)
- Coleps hirtus viridis (Ehrenberg, 1831) (ref. ID; 4613 redescribed paper)
Syn; Coleps viridis Ehrenberg, 1831 (ref. ID; 4613) - Coleps incurvus Ehrenberg, 1833 (ref. ID; 1619, 2141, 3540) or 1841 (ref. ID; 3115) reported author and year? (ref. ID; 4611, 4613)
See; Coleps hirtus (ref. ID; 3115) - Coleps inermis Perty, 1852(?)
See; Enchelys pellucida Eberhard, 1862 (ref. ID; 3540) - Coleps inermis Perty, 1858(?)
See; Coleps striatus Smith, 1897 (ref. ID; 1619, 3540) - Coleps kenti Bathia, 1922
See; Coleps striatus Smith, 1897 (ref. ID; 1619, 3540) - Coleps niger Lepsi, 1965 (ref. ID; 2141)
- Coleps nolandi Kahl, 1930 (ref. ID; 1629, 2100, 2141, 3540, 4611, 4613, 7759) reported year? (ref. ID; 3698) reported author and year? (ref. ID; 4266)
- Coleps octospinus Noland, 1925 (ref. ID; 1308, 1619, 2141, 3540) reported year? (ref. ID; 1618)
- Coleps pulcher Spiegel, 1926 (ref. ID; 1619, 1890, 2117, 2141, 2249, 3119, 3540, 3690)
- Coleps quadrispinus Foissner (ref. ID; 3698, 4613)
- Coleps remanei Kahl, 1933 (ref. ID; 1620, 2141)
- Coleps similis Kahl, 1933 (ref. ID; 1620, 2117, 2141, 3119)
- Coleps spetai Foissner, 1984 (ref. ID; 4611, 4613) reported year? (ref. ID; 6141) reported author and year? (ref. ID; 1629, 7040)
- Coleps spiralis Noland, 1937 (ref. ID; 1618, 2141, 3119)
- Coleps striatus Smith, 1817 (ref. ID; 2141) or 1897 (ref. ID; 1619, 3540)
Syn; Coleps inermis Perty, 1858(?) (ref. ID; 1619, 3540); Coleps kenti Bathia, 1922 (ref. ID; 1619, 3540) - Coleps tesselata Kahl (ref. ID; 2117)
- Coleps tesselatus Kahl, 1930 (ref. ID; 1619, 2100, 2141, 2316, 3119, 3540) reported year? (ref. ID; 3690, 3771, 3846)
Syn; Cercaria hirta O.F. Muller? (ref. ID; 3540) - Coleps trichotus Savi, 1913 (ref. ID; 1619, 2141, 3540)
- Coleps uncinatus Claparede & Lachmann, 1859 (ref. ID; 1619, 2141, 3540)
- Coleps uncinatus Roux, 1889
See; Coleps amphacanthus Ehrenberg, 1833 (ref. ID; 1619, 3540) - Coleps vesiculosus Lepsi, 1965 (ref. ID; 2141)
- Coleps viridis Ehrenberg, 1831 (ref. ID; 4613) or 1838 (ref. ID; 1619, 3115, 3540)
See; Coleptus hirtus (ref. ID; 1619, 3115, 3540), Coleps hirtus viridis (ref. ID; 4613)
Coleps amphacanthus Ehrenberg, 1833 (ref. ID; 1335, 1619, 2141, 3540, 4266) reported year? (ref. ID; 4613) reported author and year? (ref. ID; 191, 4611)
Synonym
Coleps uncinatus Roux, 1889 (ref. ID; 1619, 3540)Descriptions
Stomatogenesis: Coleps amphacanthus like other species of this genus has five girdles of skeletal plates. These plates are made of calcified organo-phosphates and are located in the pellicular alveoli. Stomatogenesis starts in the equatorial region of the cell between the anterior and posterior main plates. It is the anterior parts of only four kineties that are involved in the kinetosome proliferation for the buccal ciliature of the opisthe. The entire process of cell division takes about one hour and can be subdivided in seven stages characterized by the following criteria. (ref. ID; 4266)- Stage 1. Kinetosome proliferation is seen in all somatic kineties, but most intensively in a particular kinety, named K1 hereafter, which is one of the four stomatogenic kineties that are involved in the formation of the oral ciliature of the future opisthe.
- Stage 2. A small zone free of kinetosomes appears between the future proter and opisthe, kinetofragment 1 is composed of about 20 pairs of kinetosomes. The axis of these dikinetids is oriented perpendicular to the axis of the kinety. Due to the more intense proliferation in this kinetofragment, the zone free of kinetosomes is shifted further towards the anterior half of the cell.
- Stage 3. The anterior end of kineties Kn, Kn-1, and Kn-2 have transformed into kinetofragments too, each composed of three to five paired kinetosomes with their axes still parallel to the longitudinal axes of their respective stomatogenic kineties.
- Stage 4. Kinetosome proliferation for the buccal anlage of the future opisthe is finished. Kinetofragment 1 has changed its position compared to the orientation of the somatic kineties. It has turned slightly towards the right side of the cell. The individual dikinetids of kinetofragment n, n-1, and n-2 have turned to the left and thus changed their orientation with respect to the long axis of the kinetofragments; they are now lying at right angles to the long axes of the latter.
- Stage 5. Kinetofragment 1 is now lying obliquely between proter and opisthe, or more precisely, it is positioned in front of the anterior end of the stomatic kineties K1 to K5 of the future opisthe. The kinetofragments n, n-1, and n-2 have separated a little from their respective kineties.
- Stage 6. Kinetofragment 1 has performed a semicircular movement and lies now in front of some 8-10 somatic kineties. The three kinetofragments on the left side of the buccal anlage have separated more distinctively from their stomatogenic kineties and are now oriented slightly oblique to the latter.
- Stage 7. The dikinetids of kinetofragment 1 have moved around the presumptive oral opening of the opisthe. Each dikinetid has thereby changed its orientation so that its axis is at a right angle to the somatic kineties.
The light microscopical observation of stomatogenesis in Coleps amphacanthus clearly shows that the paroral dikinetids which surround almost the entire oral opening are derived from one single somatic kinety, K1, whereas the three adoral organelles, formerly known as "brosse" originate from three additional somatic kineties, Kn, Kn-1, and Kn-2. (ref. ID; 4266)
Measurements
70-90 um. (ref. ID; 2141)Coleps bicuspis Noland, 1925 (ref. ID; 1619, 2141, 2245, 3540) reported year? (ref. ID; 1618, 4613) reported author and year? (ref. ID; 4266)
Descriptions
16 rows of platelets; 2 posterior processes. (ref. ID; 1618)Measurements
About 55 um long. (ref. ID; 1618, 2141)Coleps dentatus Lepsi, 1965 (ref. ID; 2141)
Measurements
60 um?. (ref. ID; 2141)Coleps elongatus Ehrenberg, 1830 (ref. ID; 1619, 2100, 2141, 3540) or 1831 (ref. ID; 4613) reported year? (ref. ID; 1618, 3690) reported author and year? (ref. ID; 4611)
Descriptions
Slender; about 13 rows (Noland, 1925) or 14-17 rows (Kahl) of platelets; 3 posterior processes. (ref. ID; 1618)Measurements
40-55 um long. (ref. ID; 1618)65-80 um. (ref. ID; 2141)
Coleps heteracanthus Noland, 1937 (ref. ID; 2141) reported year? (ref. ID; 1618)
Descriptions
Anterior processes only on one side; posterior spines; caudal cilium; salt water. (ref. ID; 1618)Measurements
About 90 by 35 um. (ref. ID; 1618)90 um. (ref. ID; 2141)
Coleps hirtus O.F. Muller, 1786 (ref. ID; 4730) reported year? (ref. ID; 1335, 1618), (O.F. Muller, 1786) Nitzsch, 1827 (ref. ID; 4611) or Nitzsch, 1817 (ref. ID; 1219, 1308, 1619, 1879, 1890, 2100, 2141, 2245, 3115, 3540) reported year? (ref. ID; 3342, 3690) reported author and year? (ref. ID; 191, 5388)
Synonym
Cercaria hirta Nitsch, 1817 (ref. ID; 4730); Cercaria hirta O.F. Muller, 1786 (ref. ID; 1619, 3115, 3540, 4611, 4730); Coleps incurvus Ehrenberg, 1841 (ref. ID; 3115); Coleps viridis Ehrenberg, 1838 (ref. ID; 1619, 3115, 3540); Dictiocoleps hirtus Diesign, 1866 (ref. ID; 3115); Vorticella punctata Abildgaard, 1793 (ref. ID; 3540)Descriptions
Two main groups of plates with 4 meridional rows of "windows" each; 15-20 longitudinal rows of platelets; mouth at the anterior pole, surround by special platelets; 3 spinous processes at the posterior end; uniform ciliation over the whole body except for 1 long caudal cilium; a spherical macronucleus; contractile vacuole near the posterior end. Other species of the genus Coleps may show very similar features; in doubtful cases, make sure of the construction of the main plates and caudal spines. (ref. ID; 1219)Fifteen to twenty rows of platelets; three posterior processes. (ref. ID; 1618)
Measurements
Length 55-65 um. (ref. ID; 1219, 2141)40-65 um long. (ref. ID; 1618)
47-50x21-24 um. (ref. ID; 3342)
Coleps hirtus hirtus (O.F. Muller, 1786) Nitzsch, 1827 (ref. ID; 4613 rediscribed paper)
Synonym
Cercaria hirta O.F. Muller, 1786 (ref. ID; 4613)Descriptions
1) Size in vivo about 40-65x18-35 um. 2) Body covered in calcified armoured plates, usually barrel-shaped, rarely cylindrical. Armour consist of 6 plate rings (from anterior): circumoral plates, anterior collateral and main plates, posterior main and collateral plates, and circumanal plates. Each ring composed of 12-20, usually 15 or 16 plates. Collateral armour plates each with 2, main plates each with 4 windows, in total thus 12 windows per plate row. Windows pretzel (=8) shaped, ridge at left plate margin finely crenellated, never wing-like as in C. spetai, teeth of right plate margin alternately arranged to windows. One large spine at an anterior collateral plate and three or four prominent spines at rear end. 3) Macronucleus globular, near mid-body. 4) Contractile vacuole in posterior end. 5) Extrusomes (toxicysts) difficult to recognize in vivo. Cells, especially armour, usually brown. No symbiotic green algae. 6) 12-20, usually 15 or 16 meridional ciliary rows, each with four condensed cilia (pectinelles) at anterior end; cilia project from small, fusiform orifices between armour plates. One distinctly elongated caudal cilium. 7) Oral opening at anterior end of cell, oral basket inconspicuous. Undulating membrane circular, composed of paired basal bodies, interrupted by three tiny adoral organelles. 8) Movement without peculiarities, swims moderately rapidly. (ref. ID; 4613)Comments
Foissner et al. (1994) assigned records of green, that is, zoochlorellae-bearing C. hirtus to C. spetai. But now we resurrented C. hirtus viridis, which is very similar to C. hirtus hirtus, except for the symbiotic algae. The following species have the same type of armour plates (windows 8-shaped): Coleps hirtus viridis (green due to symbiotic algae), C. spetai (green due to symbiotic algae, more distinctly barrel-shaped, main armour plates with wing-like marginal ridge, and, most important, anterior main plates with 5 windows each), C. elongates (2 caudal cilia, 5 windows in anterior main plates), C. amphacanthus Ehrenberg (larger, more windows, several caudal cirri, bursiform), C. bicuspis Noland (Only 2 spines at rear end). Coleps nolandi has armour plates with reniform windows. (ref. ID; 4613)Coleps hirtus var. lacustris Faure-Fremiet, 1924 (ref. ID; 1619, 2141, 3540) reported author and year? (ref. ID; 3292)
Measurements
60 um. (ref. ID; 2141)Coleps hirtus var. minor Kahl, 1930 (ref. ID; 1619, 2141, 3540 original paper, 4611, 4613)
Measurements
35-45 um. (ref. ID; 2141)Coleps hirtus viridis (Ehrenberg, 1831) (ref. ID; 4613 redescribed paper)
Synonym
Coleps viridis Ehrenberg, 1831 (ref. ID; 4613)Descriptions
Size (40-50x25-35 um, n=4, original data, in vivo), shape, armour, macronucleus, contractile vacuole, and somatic and oral ciliary pattern as in Coleps hirtus hirtus. The only difference is the occurrence of symbiotic green algae (few to many, occasionally even lacking, 5-6 um across, without eyespot), which should not be confused with ingested green algae. Coleps spetai is also green due to symbiotic algae, but its main plate have wing-like ridges, and, most important, each anterior main plate has 5 windows (against 4 in C. hirtus viridis and C. hirtus hirtus). Four windows in each anterior margin plate and the symbiotic green algae are thus key characters for identification. (ref. ID; 4613)Comments
Previously we assigned record of green Coleps hirtus populations to C. spetai (Foissner et al. 1994), another symbiotic algae bearing species. A detailed reinvestigation of a green C. hirtus population did not show any significant morphological differences to aposymbiotic C. hirtus populations. The "green C. hirtus" is very likely C. viridis of Ehrenberg, which was synonymized with C. hirtus by Kahl (1930). However, symbiotic and aposymbiotic populations have a different physiology and ecology, suggesting that they should be considered as subspecies. Furthermore, the symbiosis is rather stable. (ref. ID; 4613)Coleps incurvus Ehrenberg, 1833 (ref. ID; 1619, 2141, 3540) or 1841 (ref. ID; 3115) reported author and year? (ref. ID; 4611, 4613)
See
Coleps hirtus (ref. ID; 3115)Measurements
65-80 um. (ref. ID; 2141)Coleps niger Lepsi, 1965 (ref. ID; 2141)
Measurements
60 um?. (ref. ID; 2141)Coleps nolandi Kahl, 1930 (ref. ID; 1629, 2100, 2141, 3540, 4611, 4613, 7759) reported year? (ref. ID; 3698) reported author and year? (ref. ID; 4266)
Descriptions
Plate morphology by SEM: Each circular girdle consists of approximately 12-14 identical plates. The anterior and posterior secondary plates as well as the posterior main plates of the investigated strain of C. nolandi are of equal size while the anterior main plates are longer. The plates of a girdle overlap each other, the right edge of one plate covering the left side of the neighbouring plate. The individual longitudinal plate is rectangular in outline and surrounded by a rim. Its longer sides are undulated. The plate is constructed like a slightly slanting roof with the ridge of the roof shifted a little to the right. The ridge ("Hauptleiste" Kahl, 1930) is formed to 4-6 parallel ribs. The left part of a plate which is normally covered by the adjacent plate is actually composed of two layers. These two layer. These two layers are connected by 3 resp. 4 pairs of vertical columns which project from the rim. Under each of these pairs of columns a single process protrudes to the inside of the cell. Between two processes there is an indentation in the lower rim to make room for a cilium. To investigate the lower layer of the plates, cells were arbitrarily fractured using a freeze-etching technique modified for SEM. This technique allows the observation of chemically untreated cells with their plates still surrounded by the alveolar membranes. The lower layer has 3 (in longer plates 4) roughly hemispherical elevations with numerous holes, the two marginal elevations being about half the size of the middle ones. The apex of these elevations fuses with the covering layer. To the right of the elevations there is a longitudinal groove exactly underneath the longitudinal ridge in the upper layer. It is along this line that the two layers show the greatest distance from each other. Further to the right two layers merge into a single one. In situ this single layered part covers the neighbouring plate. (ref. ID; 7759)Notes
SEM studies of the endoskeleton of Coleps have been made so far only with representatives of type 1. In this study the species with type 2 plates, C. nolandi, was investigated. In contrast to the skeletal plates of C. hirtus and C. amphacanthus those of C. nolandi are composed of two layers which join at various points. The lower layer of the plate, which resembles a type 1 plate in its fenestrated elevations, is covered by a roof-like upper layer which shows a mesh-like structure. Although it is not possible to detect the double layered architecture of the plates at light microscopical level, Kahl (1930) gave a detailed and almost correct Descriptions of their morphology. He pointed out that the conspicuous ridge ("Hauptleiste") at the right edge of type 1 plates is shifted towards the middle in C. nolandi plates. In C. hirtus the lateral processes ("Zahne") alternate with the subdivided elevations, while in C. nolandi they are placed in front of the undivided elevations. He assumed the perforated elevations ("Fenster") in C. nolandi to be vaulted. Kahl described the right side of a plate to be rather fragile, this is identical with the single layered part which covers the adjacent plate. The schematic drawings of the plated given by Kahl represent a two-dimensional object. The SEM investigation reveals principal differences in plate morphology between type 1 and type 2. While the type 1 plates are single layered with hollow spaces in all thicker parts, e.g. lateral processes, the longitudinal ridge and the surrounding rim, the double layered type 2 plates show hollow space between upper and lower layer. (ref. ID; 7759)Measurements
50-65 um. (ref. ID; 2141)Coleps octospinus Noland, 1925 (ref. ID; 1308, 1619, 2141, 3540) reported year? (ref. ID; 1618)
Descriptions
8 posterior spines; about 24 rows of platelets; Geiman (1931) found this organism in an acid marsh pond and noted variation in number and location of accessory spines. (ref. ID; 1618)Measurements
80-110 um long. (ref. ID; 1618)100-110 um. (ref. ID; 2141)
Coleps pulcher Spiegel, 1926 (ref. ID; 1619, 1890, 2117, 2141, 2249, 3119, 3540, 3690)
Descriptions
In marine. (ref. ID; 2249)Measurements
100 um. (ref. ID; 2141)Coleps remanei Kahl, 1933 (ref. ID; 1620, 2141)
Measurements
50-70 um. (ref. ID; 2141)Coleps similis Kahl, 1933 (ref. ID; 1620, 2117, 2141, 3119)
Measurements
200-250 um. (ref. ID; 2141)Coleps spiralis Noland, 1937 (ref. ID; 1618, 2141, 3119)
Descriptions
About 23 longitudinal rows of platelets slightly spirally twisted; posterior spines drawn together; a long caudal cilium; salt water. (ref. ID; 1618)Measurements
About 50 um long. (ref. ID; 1618)50 um. (ref. ID; 2141)