Askenasia
Askenasia Blochmann, 1895 (ref. ID; 2013)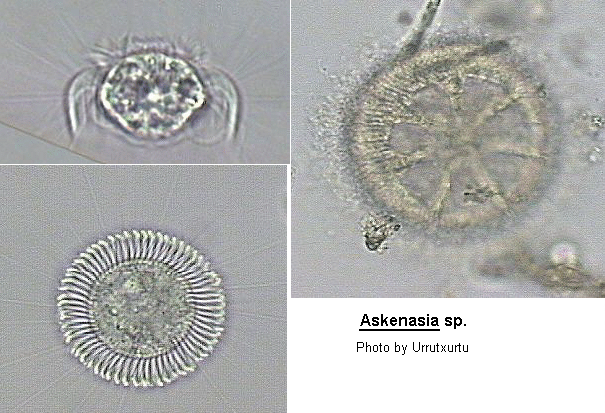
Class Kinetofragminophora De Puytorac et al., 1974: Subclass Gymnostomata Butschli, 1889: Order Haptorida Corliss, 1974: Family Didiniidae Poche, 1913 (ref. ID; 4798) [ref. ID; 2013]
Body shape approximately pyriform, with larger, rounded posterior region and a smaller domed anterior region. Oral aperture apically situated, cytopharynx supported by trichites. There are 2 ciliary bands set close together in the anterior half of the cell. The anterior most band is composed of short rows of short cilia projecting forward and the posterior most band of longer rows of long cilia projecting backwards. In some species there is a circle of bristles (cirri) immediately behind the posterior ciliary band, these project stiffly out radially. In one species the posterior half of the body is sparsely ciliated. Macronucleus spherical to ovoid. Contractile vacuole terminal.
Quote; Colin R. Curds "British and other freshwater ciliated protozoa Part I Ciliophora: Kinetofragminophora" Cambridge University Press, 1982 (ref. ID; 2013) [ref. ID; 4352]
Nomenclature; The genus name, Askenasia, is a junior synonym established by Blochmann (1895). It has to be retained in accordance with the IRZN, article 23, because the senior name, Stephanidina, is considered as nomen oblitum (Foissner 1987). There is much confusion concerning the synonymy and nomenclature of the type species. A. volvox. It was originally described by Eichward (1852) as Trichodina volvox. Furthermore, he proposed to establish Stephanidina as a new genus for this species. Trichodina volvox was then transferred to the genus Halteria (Claparede & Lachmann, 1859). At the end of the 19th century Blochmann (1895) published. A. elegans as a new genus and species, unfortunately overlooking the valid original description as well as the redescription (Claparede & Lachmann 1859). His name is thus a synonym. Kahl (1930) recognized the synonymy of Halteria volvox and A. elegans and consequently designated Blochmann's species as A. volvox (Claparede & Lachmann, 1859), but also disregarded the original description; hence listing the wrong author and date. Finally, Dingfelder (1962) 1st mentioned the original description by Eichwald, but incorrectly stated "Blochmann, 1895" as the 1st revising author. However, Kahl is the 1st revisor and the correct citation of the taxon must be "Askenasia volvox (Eichwald, 1852) Kahl, 1930" (Foissner 1987). (ref. ID; 4352)
General morphology and ecology of Askenasia; The size ranges from 27-60 um in length and from 22-65 um in width, with length and width usually approximately equal. The body has an ovoid to diamond-shaped outline and can be divided into an anterior cone-shaped "rostrum" and a posterior hemispherical portion, both circular in transverse section. The cell equator represents the transition between the anterior and the posterior portion, which does not always correspond to the geometrical median plane of the cell. The shape of the rostrum varies from sharply truncated through arched to tapered. The posterior portion is broadly rounded to bluntly pointed. A single globular to rope-like macronucleus is situated in the posterior portion or just below the cell equator. Occasionally, specimens (presumably postconjugates) were found with up to 10 globular segments. Except in A. stellaris, which possesses two macronuclear segments and two micronulei, a single spherical micronucleus is attached to the macronucleus in variable position. One, two or four contractile vacuoles discharge by conspicuous excretory pores subequatorially. The vacuole is either a simple blister or is surrounded by collecting vesicles during diastole. Multiple vacuoles are diametrically opposed and function alternately. No data are known on the cytoproct. Three species have an enigmatic crescent-shaped vacuole on the rear (we suggest the term "terminal vacuole") which is hyaline and often disappears in vivo. Protargol slides show that it adheres to the posteriormost transverse bristle row. The bristles are bent posteriad forming a basket-shaped structure which enclose the vacuole. Whether the vacuole contains an air-bubble or some sort of liquid is not known. Nonetheless, we suggest that it has a special function in the planktonic mode of life, possibly increasing buoyancy. Mesodinium pulex Claparede & Lachmann, 1858, was observed to also have such a terminal vacuole (Borror 1963). The pellicle is ribbed (seen only in optical transverse section, with the cilia extending between the ribs) and fragile. Coverslip pressure usually causes immediate lysis. Extrusomes occur in several approximately pin-like types which are packed in bundles surrounding the cytopharynx, but often a few are scattered throughout the cytoplasm. Shape and arrangement of the extrusomes represent important species characters. Askenasia species feed on algae and other protozoa (Dingfelder 1962; Tamar 1973). Ingestion and defecation have, however, not been observed.
Locomotion is a highly characteristic feature in the genus. The "pectinelles" of the pre-equatorial kinety belt (PKB) beat in a rotating ring consisting of 5-16 flame-like tufts, impressively visible during the immobile period. They draw food particles toward the cytostome, while the cell rotates leisurely around the transverse axis (Tamar 1973). The lighting darts and short, spiralling jumps, which alternate with abrupt stops, are caused mainly by the bell-shaped skirt, formed from the "cirri" of the equatorial kinety belt (EKB), pushing the water backwards. It is also evident that the cirral-skirt, when immobile, keeps the cell afloat. The bristles of the subequatorial kinety belt (SKB) act as stabilizers during floating but may also play a role in forward movement by contracting (Penard 1922; Tamar 1973). Three belts, of differently arrayed cilia, mantle the anterior portion. Each belt is composed of (47-)60-70 roughly longitudinal kineties and separated from its neighbor by a small gap. The 8-15 um long pectinelles (P) originate from the pre-equatorial kinety belt (PKB), whose kineties are composed of single files of basal bodies and are interconnected by transverse fibers. The equatorial kinety belt (EKB) bears the rigid, sickle-shaped, about 20-30 um long "cirri", which form a skirt around the posterior half of the cell. Each cirrus is composed of a double row of basal bodies arranged in zigzags and interconnected by an argyrophilic fiber. The subequatorial kinety belt (SKB) contains 20-60 um long, flexible bristles. Each kinety is obliquely oriented to the longitudinal cell axis and comprises of three basal bodies, usually interconnected by transverse fibers. The posterior portion shows oblique single files of non-ciliated granules (PG, smaller than basal bodies) which usually lie in the center of weakly impregnated square structures, producing more or less regular lattice. However, A. chlorelligera differs noticeably in the absence of these linear files an din possessing a network of polygons resembling honeycombs and enclosing single or paired granules. This network corresponds closely to the silverline system. The cytostome-cytopharyngeal complex is located in the long axis of the cell and is usually equipped with conspicuous extrusomes. The cytostome, lying somewhat depressed in the center of the anterior pole (oral area), is round to slit-like (Tamar 1973). The oral area is encircled by a remarkable "circumoral wreath of unciliated, paired granules" (CWG), which has a circular or more or less cloverleaf-shape. Each pair usually comprises of a larger posterior and a smaller anterior granule. Both are slightly inclined to the cell's long axis and are of equal size in A. chlorelligera. To date we are unable to decide whether the granules are basal bodies or mucocysts or something else, and whether the smaller satellites are parasomal sacs or not. Accordingly, we prefer the term "circumoral wreath" to "circumoral kinety". The granule pairs are usually associated with weakly impregnating fibers (longitudinal fibers, LF), which course posteriorly to the pre-equatorial kinety belt (PKB) and anteriorly to the cytostome (transverse fibers, TF). The CWG is an important diagnostic feature at species level. There is now strong evidence (cp. A. volvox) that the apical star-like figure in A. stellaris, which Kahl assumes to be pharyngeal wrinkles, is also a CWG. The silver line system is widely meshed, both in the posterior portion and in the oral area, whereas the middle cell are shows circumferential bands of rectangles, and subjacently, honeycombed structures. Bipartition by transverse fission (homothetogenic fission, Tamar 1973) has not yet been studied in detail. It begins in the equatorial kinety belt with a constriction (Penard 1922). The related genus Mesodinium shows no constriction during bipartition (Tamar 1987). The macronucleus appears dumbbell-shaped in the late fission stages. The proter reorganizes the equatorial kinety belt, the opisthe the pre-equatorial kinety belt, while the reorganization of the bristles has not yet been observed. The pectinelles are active before the dividers separate. Conjugation occurs by fusion at the anterior end. During conjugation the pectinelles do not cease beating. The fragmented macronuclear material is present in the anterior portions of the conjugants (Penard 1922; Tamar 1973). The resting cyst is known only for A. volvox (Penard 1922); it consists of two shell-like parts which are equipped with spines.
Despite extensive biogeographic data, little information on the ecology is available. Askenasia inhabits limnetic as well as marine ecosystems, preferentially the plankton. It was recorded from many localities in the holarctic region (see A. volvox), but is still not documented from the southern hemisphere. In freshwater ecosystems Askenasia is usually found in less polluted habitats, achieving highest densities in spring and fall when algae are abundant (Kahl 1930, Nauwerck 1963). Therefore we conclude that Askenasia is euplanktonic and generally prefers oligo- to mesotrophic conditions. Further ecological investigations on Askenasia promise to be worthwhile. (ref. ID; 4352)
Taxonomic changes; Askenasia comprises four valid species in our opinion. Two other nominal species, A. faurei and A. humilis, ought to be transferred to Cyclotrichium.
Cyclotrichium faurei (Kahl, 1930) nov. comb. was 1st described as Askenasia elegans (Faure-Fremiet 1924). Kahl (1930) later classified it, in agreement with Faure-Fremiet, as a new species, A. faurei. Askenasia faurei, however, lacks bristles, has a ciliated posterior portion and a single contractile vacuole with collecting canals in the rear. These characters are typical for Cyclotrichium (Faure-Fremiet 1924; Kahl 1930; Wilbert 1986). Cyclotrichium faurei, however, requires redescription based on silver impregnation. It is a rare species reported only twice since the original description (Bernerth 1982; Burkovsky 1971).
Cyclotrichium humilis (Gajewskaja, 1928) nov. comb. is a bizarre-shaped species, which has, like C. faurei, two kinety belts and fringe-like cilia associated with the equatorial kinety belt. It also requires redescription, and has not been recorded since the original description. The type species of Cyclotrichium has only a single anterior kinety belt (Kahl 1930). Therefore C. faurei and C. humilis, each with two kinety belts, may be transferred to a new genus in the future. (ref. ID; 4352)
Improved diagnosis of Askenasia; Mesodiniidae with: (i) one circumoral wreath of argyrophilic granules; (ii) three circumferential, equatorial kinety belts (kineties of pre-equatorial belt composed of single files of basal bodies, kineties of equatorial belt composed of double rows of basal bodies arranged in a zigzag, kineties of subequatorial belt composed of three basal bodies each); and (iii) more or less needle or pi-shaped extrusomes, usually surrounding cytopharynx. Free-living, carnivorous and algivorous, freshwater and marine. (ref. ID; 4352)
Remarks; According to Corliss (1979), Askenasia belongs to the class Kinetofragminophora, subclass Gymnostomata, order Haptorida, family Didiniidae. Our investigations show that Askenasia lacks important haptorian characteristics: the distinct rhabdos, the dorsal brush, and the tightly meshed silverline system (Corliss 1979; Foissner & Foissner 1988). Thus, we transfer Askenasia to the order Cyclotrichida Jankowski, 1980, family Mesodiniidae Jankowski, 1980, whose key features readily match those of Askenasia (Foissner & Foissner 1988). However, the Cyclotrichida themselves are in urgent need of investigation and revision under silver impregnation techniques. The Cyclotrichida may possibly belong not to the subclass Haptoria but to another (Foissner & Foissner 1988). (ref. ID; 4352)
Type species; Askenasia volvox (Eichwald, 1852) Kahl, 1930 (ref. ID; 4352)
- Askenasia acrostomia Krainer & Foissner, 1990 (ref. ID; 4352 original paper, 4612, 4613) reproted author and year? (ref. ID; 7040)
- Askenasia chlorelligera Krainer & Foissner, 1990 (ref. ID; 4352 original paper, 4613) reproted author and year? (ref. ID; 7040)
- Askenasia elegans Blochmann
See; Askenasia volvox (ref. ID; 2245, 3540, 4352, 4613) - Askenasia elegans Faure
See; Askenasia faurei Kahl, 1930 (ref. ID; 3540) - Askenasia faurei Kahl, 1930 (ref. ID; 3540 original paper) reported year? (ref. ID; 1618, 1619)
Syn; Askenasia elegans Faure, 1924 (ref. ID; 3540) - Askenasia stellaris Leegaard, 1920 (ref. ID; 1619, 3540) or (Leegaard, 1920) Kahl, 1930 (ref. ID; 4352)
Syn; Lohmanniella stellaris Leegaard, 1920 (ref. ID; 4352) - Askenasia volvox Claparede & Lachmann, 1858 (ref. ID; 1335, 1619, 1620, 1629, 2245, 3540, 4671, 4798) reported year? (ref. ID; 1219) reported author and year? (ref. ID; 3954)
See; Halteria volvox (ref. ID; 3389)
Syn; Askenasia elegans Blochmann (ref. ID; 2245, 3540); Mesodinium recurva Kellicott, 1885 (ref. ID; 1619, 3540) - Askenasia volvox (Eichwald, 1852) Kahl, 1930 (ref. ID; 4352, 4488, 4612, 4613)
Syn; Askenasia elegans Blochmann, 1895 (ref. ID; 4352, 4613); Halteria volvox Claparede & Lachmann, 1858 (ref. ID; 4352); Trichodina volvox Eichward, 1852 (ref. ID; 4352, 4612, 4613)
Askenasia acrostomia Krainer & Foissner, 1990 (ref. ID; 4352 original paper, 4612, 4613) reproted author and year? (ref. ID; 7040)
Diagnosis
Size in vivo 40-50x30-40 um. Circumoral wreath of granules circular. Macronucleus horseshoe-shaped to rope-like. Extrusomes needle-like with pointed ends. Four contractile vacuoles. (ref. ID; 4352)Descriptions
Outline approximately rhomboid with distinct cell equator. Anterior portion tapering, constricted at level of oral area, where a single extrusome bundle protrudes. Well-fed specimens with globular posterior portion. Macronucleus with chromatin bodies of various shaped and sizes. Several specimens observed with two sausage-like fragments (resembling A. stellaris). Micronucleus usually located at end of a macronuclear arm. Four contractile vacuoles diametrically arranged in posterior 3rd of cell, discharging by short excretory canals about 1 um in length. Bases of excretory canals framed by argyrophilic fibers. Extrusomes about 23 um long, extending at least 2/3 of cell length posteriorly from apex. They usually discharge on fixation, forming tangles of threads; in protargol slides having knobbed posterior ends not discernible in vivo. Furthermore, impregnations suggest presence of a 2nd, shorter type. We have not, however, ascertained it is vivo. Cytoplasm of starved specimens, colorless in posterior portion some greasily shining inclusion. Feeds on algae and ciliates. Terminal vacuole discernible in vivo. The EKB-kineties are approximately two kinety widths apart with basal bodies loosely spaced. The SKB-kineties are interconnected by transverse fibers. Granule pairs of CWG do not show fibers under impregnation method. (ref. ID; 4352)Etymology
"acrostomia" refers to the tapered rostrum. (ref. ID; 4352)Type location
Excavated groundwater ponds near Graz (Styria, Austria). (ref. ID; 4352)Askenasia chlorelligera Krainer & Foissner, 1990 (ref. ID; 4352 original paper, 4613) reproted author and year? (ref. ID; 7040)
Diagnosis
Size in vivo 30-40x25-45 um. Circumoral wreath of granules roughly pentagonal. Macronucleus spheroidal to ellipsoid (3:2). Two types of pin-shaped extrusomes. Single contractile vacuole. (ref. ID; 4352)Descriptions
Rostrum truncated, crown-like structure comprised of 5-6 bumps apically, produced by protruding bundles of shorter extrusome type. "Crown" resembles cross-section of apple-core. Macronucleus with special chromatin bodies of various sizes. Several specimens with two spheroidal fragments. Excretory canal of contractile vacuole about 2 um long. Short extrusomes about 10 um, knobbed proximally, packed in posteriorly slightly diverging, in cross-section elliptical bundles. Long extrusomes about 18 um, slightly curved, extending half cell length posteriorly from crown, forming a basket in the center of the cell. Extrusome tips impregnated conspicuously with protargol. Cytoplasm contains about 20-30 Chlorella-algae and several green algae seemingly in terms of several characters, belonging to Siderocelis. Algae give cell a grass-green color. Terminal vacuole discernible in vivo. The EKB-kineties are about one kinety width apart, basal bodies comparatively loosely spaced. The EKB-kineties have short trunks of 5-8 basal bodies at the posterior end. The SKB-kineties interconnect by transverse fibers. The posterior hemisphere shows polygonal, argyrophilic pattern. At the cell's equator there is a circumferential belt of rectangular structures linking polygons with EKB-kineties. The CWG comprises of paired granules of equal size, resembling a haptorid oral dikinetid, encircling extrusome bumps. (ref. ID; 4352)Remarks
This is obviously the green, pelagic A. volvox which Kahl (1935) found in the plankton of several freshwater ponds near Hamburg, although already assuming into be a new species. (ref. ID; 4352)Etymology
"chlorelligera" refers to the symbiotic green algae. (ref. ID; 4352)Type location
Excavated groundwater ponds near Graz (Styria, Austria). (ref. ID; 4352)Askenasia faurei Kahl, 1930 (ref. ID; 3540 original paper) reported year? (ref. ID; 1618, 1619)
Synonym
Askenasia elegans Faure, 1924 (ref. ID; 3540)Descriptions
Body oval, anterior end broadly rounded; posterior region conical; pectinelles about 13 um long; the second band (10 um) of long cilia; an ellipsoid macronucleus; a micronucleus; fresh water. (ref. ID; 1618)Measurements
Body about 58-60 um long. (ref. ID; 1618)Askenasia stellaris Leegaard, 1920 (ref. ID; 1619, 3540) or (Leegaard, 1920) Kahl, 1930 (ref. ID; 4352)
Synonym
Lohmanniella stellaris Leegaard, 1920 (ref. ID; 4352)Descriptions
Lentil-shaped to somewhat flattened globular. Rostrum sharply truncated, posterior portion dish-shaped to bluntly pointed. Oral area shows star-like figure with 6-8 lobes encircling cytostome, very likely a CWG. Two sausage to worm-shaped macronuclear segments, eccentrically located in cell equator, each part enclosed in special membrane (Borror 1963), size of each part approximately 17x7 um (Borror 1963). Two micronuclei, about 4 um in diameter each, closely attached to inner concave portion of macronuclear segments. Two contractile vacuoles with collecting vesicles located equidistantly from macronuclei. Extrusomes ("trichites") rod-shaped, about 20 um in length, arranged in two whorls of six bundles with 12-20 extrusomes each, protruding from rostrum. Feeds on cocci and algae. Kinety belts comprise approximately 70 kineties. Pectinelles beat in 6-16 tufts (Borror 1963; Kahl 1931). Bristles from 20-60 um in length (Borror 1963; Kahl 1931). (ref. ID; 4352)Remarks
This marine species was originally described under the name Lohmanniella stellaris. It was transferred by Kahl (1930) to the genus Askenasia. Askenasia stellaris is the sole marine species described. It is well characterized by its nuclear and excretory apparatus. However, redescription with silver impregnation techniques is required. (ref. ID; 4352)Measurements
Size in vivo 24-50x29-65 um (Borror 1963; Kahl 1931; Leegaard 1920). (ref. ID; 4352)Askenasia volvox (Eichwald, 1852) Kahl, 1930 (ref. ID; 4352, 4488, 4612, 4613)
Synonym
Askenasia elegans Blochmann, 1895 (ref. ID; 4352, 4613); Halteria volvox Claparede & Lachmann, 1858 (ref. ID; 4352); Trichodina volvox Eichward, 1852 (ref. ID; 4352, 4612, 4613)Surprisingly, we did not find a form which correspond exactly to any of the available descriptions of A. volvox. We observed, however, a rather similar species which differs only by more C-shaped macronucleus and the conspicuous excretory canal. Initially, we decided that it should be described as a new species; however, this view was discouraged by one of the reviewers. Thus, we withdraw our decision by separate the descriptions. Future research well decide whether this was wise or not. (ref. ID; 4352)
Descriptions
- From literature: Shape ovoid to pyriform. Macronucleus globular to ellipsoid (12-14.5x9.5-11.5 um; Tamar 1973), contains small chromatin globules. Micronucleus 3-4 um in diameter, very pale. Single contractile vacuole with collecting vesicles (Tamar 1973). Terminal vacuole present, misinterpreted by Penard (1922) as artifact of coverslip pressure. Pellicle fairly fragile, flexile (Tamar 1973). About 10 thin, needle-shaped conically bundled extrusomes ("trichites"), approximately 10-12 um in length (Penard 1922; Tamar 1973). Cytoplasm hyaline to vesicular, contains globules and tiny granules. In addition, spheres 2-3 um in diameter, dark oval particles, irregular masses as well as yellow and greenish yellow inclusions (presumably Bodo-remnants) are recognizable. Five to 12 pectinellar tufts (Kahl 1930; Penard 1922; Tamar 1973). Each kinety belt composed of about 60 kineties (Blochmann 1895; Kahl 1931; Penard 1922); according to Tamar (1973) only 47-48. Pre-equatorial belt kineties composed of about 10 basal bodies each, their cilia about 8-13 um long (Kahl 1930; Tamar 1973). Cirri composed of two adjacent rows of 14-17 basal bodies each, cilia approximately 20 um long. Bristles 30-40 um in length (Kahl 1930; Tamar 1973). (ref. ID; 4352)
- Austrian population: Outline plump-pyriform. Diameter, including cirri, about 60 um. Macronucleus C-shaped, usually with pointed ends. Chromatin bodies of various size. Some specimens with 2-5 spherical fragments. Single contractile vacuole with conspicuous excretory canal 4-6 um in length, at canal base about 3 um, at canal pore about 2 um in diameter. Extrusomes 15-22 um long (varying length by virtue of extruded extrusomes?), thickened proximally, arranged around cytopharynx, imitating CWG-pattern. Cytoplasm contains algae and cytopharyngeal baskets of ingested ciliates (presumably Urotricha spp.). Basal bodies of PKB-kineties ellipsoid, presumably because of impregnated bases of transverse fibers. The EKB-kineties slightly sigmoid, about one kinety-width apart, basal bodies densely spaced. The SKB-kineties almost rectangularly bent, each anteriormost basal body markedly smaller; no transverse fibers recognizable. Granule pairs of CWG form 6-12 arched-shaped lobes, resembling a compounded three- or four-leaf clover. Transverse and longitudinal fibers form an elaborate pattern of inverted V's, their tips pointing toward cytostome. (ref. ID; 4352)
Measurements
- From literature: Size in vivo 29-42(-50)x22-27(-50) um (Blochmann 1895; Kahl 1930; Penard 1922; Tamar 1973). (ref. ID; 4352)
- Austrian population: Size in vivo 35-45x30-45 um. (ref. ID; 4352)